Physiological responses to a short-term, environmentally realistic, acute heat stress in Atlantic salmon, Salmo salar
Abstract
Atlantic salmon populations are declining, and warming river temperatures in the summer months are thought to be a significant contributing factor. We describe the time course of cellular and metabolic responses to an ecologically relevant short-term thermal cycle in juvenile Atlantic salmon. We then examined whether this heat event would affect tolerance to a subsequent heat shock in terms of critical thermal maximum (CTmax). Fish induced heat shock protein 70 in red blood cells, heart, liver, and red and white muscle; whole blood glucose and lactate transiently increased during the heat cycle. In contrast, we observed no significant effect of a prior heat shock on CTmax. The CTmax was positively correlated with Fulton’s condition factor suggesting that fish with greater energy reserves are more thermally tolerant. Atlantic salmon activate cellular protection pathways in response to a single thermal cycle and appear to cope with this short-term, ∼1 d heat shock, but this challenge may compromise the ability to cope with subsequent heat events.
Introduction
Temperature remains one of the most significant abiotic factors affecting the biology of fishes (Fry 1958; Hochachka and Somero 2002; Currie and Schulte 2014) primarily because their body temperatures change concomitantly and rapidly with ambient temperatures (Stevens and Sutterlin 1976). High water temperatures in the summer months have been implicated in the decline of the Atlantic salmon (COSEWIC 2010) in Canada, one of the country’s most important commercial and recreational fishes. For example, the Miramichi River in New Brunswick, Canada, home to one of North America’s largest Atlantic salmon runs, is reaching exceptionally high water temperatures during the summer months (e.g., 27–30 °C; Caissie et al. 2014), exceeding the 23 °C proposed upper tolerance limit of Atlantic salmon as often as 62 d/year (Lund et al. 2002; Caissie et al. 2012). Beyond this temperature, adult Atlantic salmon have been reported to cease feeding, abandon territories, and aggregate in cool water refugia (Breau et al. 2007, 2011). Temperature changes are rapid, greater than 8 °C/d and up to 1.4 °C/h, allowing little acclimatization or recovery time (Lund et al. 2002; DFO 2012). It is important that we understand tolerance limits and the physiological changes underpinning temperatures representative of thermal events that Atlantic salmon would normally experience in the wild.
Most of our understanding regarding the effects of acute temperature change on salmonids comes from studies where fish are acclimated to one temperature and subjected to dramatic and rapid changes (i.e., over 1–2 h; e.g., DuBeau et al. 1998; Currie et al. 2000; Galloway and Kieffer 2003; Fowler et al. 2009). Although such studies provide important mechanistic insight into the capacity of salmonids to cope with acute thermal stress, they do not readily allow extrapolation to natural conditions. Comparatively fewer studies examine acute thermal scenarios representative of field conditions (but see Threader and Houston 1983; Houston and Gingras-Bedard 1994; Mesa et al. 2002; Podrabsky and Somero 2004; Todgham et al. 2006; Narum et al. 2013; Eldridge et al. 2015 for fish examples). Recently, Tunnah et al. (2017) compared physiological responses to distinct, multiday natural diel thermal cycling scenarios in wild Atlantic salmon and found no obvious differences, concluding that cycling itself is more important than the nature of the thermal cycle (Tunnah et al. 2017). In contrast, in rainbow trout metabolic reserves are depleted in a single diel thermal cycle (16–24 °C) but recover with multiple days of cycling (Callaghan et al. 2016). One outstanding question from this collective work was the immediate physiological effects of a short-term, environmentally relevant, acute heat shock, and whether or not this exposure could influence tolerance to a later, more severe thermal stress. If we hope to forecast impacts of climate warming on fishes, we need early temporal resolution to assess physiological and cellular changes in environmentally realistic contexts.
Our first goal was to describe the responses to a single heat challenge in the lab representative of field conditions, in an effort to understand the underlying physiological responses. To this end, we exposed hatchery-reared Atlantic salmon to thermal conditions representative of a well-known Atlantic salmon-producing river. We measured key metabolites (e.g., glycogen, glucose, and lactate) in tissues before, during, and following a ∼1 d thermal challenge. Cellular and endocrine stress responses have been associated with summer field temperatures in salmonid fish (Chadwick et al. 2015); thus, we also measured tissue heat shock protein 70 (HSP70) and predicted that these variables would increase with the thermal challenge. Given that the temperatures at which reductions in whole animal function occur are highly correlated with subcellular function (Anttila et al. 2013), an understanding of the molecular-level effects of natural thermal cycles could provide insight into thermal tolerance. Our second objective was to determine if this thermal cycle, based on environmentally relevant temperatures, would affect subsequent acute thermal tolerance, as measured by the critical thermal maximum (CTmax), compared with fish maintained under control conditions. It was recently demonstrated that an appreciation of fishes’ true thermal sensitivity to climate warming requires an understanding of their upper thermal physiological limits (Sandblom et al. 2016), making CTmax a particularly important dependent variable. We predicted that acute thermal tolerance would increase in Atlantic salmon exposed to a prior high temperature event.
Materials and methods
Animals
Atlantic salmon, post-smolt, were obtained from the Mactaquac Department of Fisheries and Oceans (DFO) Fish Culture Station (French Village, New Brunswick, Canada) in July 2012. These fish were the F1 progeny of early run, wild Saint John River fish, captured at the Mactaquac dam. As such, our fish were born and reared in the hatchery where they were acclimated to freshwater, held at 10 °C (±1 °C), 92% dissolved oxygen (DO), and a natural photoperiod. Fish (n = 57) were transported to the Harold Crabtree Aqualab at Mount Allison University (New Brunswick, Canada), and divided among cylindro-conical holding tanks maintained at 12 °C (±1 °C), to obtain a stocking density of approximately 21 kg/m3 (Turnbull et al. 2005). Within 2 h of arrival, fish were treated with an acute salt bath as a preventative measure against fungus and external parasites that could result from the stress of transportation and handling. Salinity was quickly (i.e., over a few minutes) raised to 20 parts per thousand (ppt) in the holding tanks and maintained for 1 h. We then decreased salinity back to 10–12 ppt within 1 h and gradually returned fish to 0 ppt over 16 h. Atlantic salmon were maintained at a natural photoperiod with DO levels >85% and fed 3 and 5 mm commercial pellet feed (Corey Nutrition Company) ad libitum every other day. After approximately 2 weeks, when all fish had begun to eat, the temperature was increased 2 °C/d at a rate of 0.1 °C/h, from 12 to 15 °C. Fish were (mean ± standard error of the mean (SEM)) 596 g ± 15.3 g, 38.6 cm ± 0.34 cm long, with a condition factor (weight/length3) of 0.0106 ± 0.0002 and held at 15 °C (±1 °C) for a minimum of 2 weeks before experiments began. Water temperature was monitored using an iBCod temperature logger (±1 °C, 15 min interval, Alpha Mach Inc.) and DO was measured daily (7.5–10 mg/L; YSI Pro 20; Xylem Inc.).
All experiments proceeded according to the Canadian Council on Animal Care guidelines approved by the Mount Allison University Animal Care Committee (MTA Protocol 12-09).
Temperature profiles
The thermal cycle (Fig. 1) was derived from hourly temperature records obtained from the Miramichi River Environmental Assessment Committee (MREAC) in July 2010 so that we could approximate the thermal conditions (e.g., peak and rate) on a typical summer day in one of North America’s most productive Atlantic salmon rivers (DFO 2013). We used hatchery fish in this study, recognizing that these fish may not be representative of the resident wild population. The thermal cycle began at 15 °C and increased to 26 °C at a rate of 0.82 °C/h; the temperature then decreased at a rate of 0.6 °C/h to return to 15 °C over 34 h. We exposed fish to these environmentally relevant temperatures and then assessed recovery. Fish were fasted 24 h before experiments began and during the thermal cycling.
Fig. 1.
Sampling
Four fish were transferred from the holding tanks to 300 L experimental tanks 24 h before experimentation with each tank representing a sampling point. This transfer was repeated once for n = 8 for each time point. Fish were terminally sampled per treatment: prior to heating (i.e., control, 0 h); at the temperature peak (15 h); 8 h following the thermal peak (23 h); when the temperature first returned to 15 °C (recovery, 34 h); or 8 h into recovery (42 h). Individual fish were anesthetized in an aerated, buffered solution of tricaine methanesulfonate (0.25 g/L; MS-222; Sigma-Aldrich, Oakville, Ontario, Canada), measured (±1 mm), and weighed (±0.1 g). Blood (5–10 mL) was sampled from the caudal vein with a 21 gauge needle, rinsed with heparinized Cortland’s saline (in mmol/L: 124.1 NaCl; 5.1 KCl; 1.9 MgSO4; 1.5 Na2HPO4; 11.9 NaHCO3; 1.1 CaCl2; 100 units/mL heparin), and immediately placed on ice. The spinal cord was then severed, and heart (ventricle), red muscle, white muscle, and liver samples were collected and immediately flash frozen in liquid nitrogen. Blood samples were analysed for whole blood glucose (One Touch Ultra 2 meter, LifeScan, Canada) and lactate (Arkray Lactate Pro meter, Fact Canada) concentrations and then centrifuged at 4 °C at 5000 rpm for 3 min to separate red blood cells (RBC) and plasma, which were immediately flash frozen in liquid nitrogen and stored at −80 °C.
HSP70 analysis
We used immunoblotting techniques to assess the relative concentrations of HSP70 in the heart, liver, red muscle, white muscle, and RBC samples. Soluble protein was extracted from the samples as performed by LeBlanc et al. (2011) and assayed using the BioRad DC Protein Assay based on the Lowry method. HSP70 levels were determined in samples (15 μg soluble protein, within the dynamic range of our chemiluminescent detection agent) as performed by Kolhatkar et al. (2014). Each gel contained the same Atlantic salmon RBC sample to express all other samples relative to this internal standard and to allow comparison across gels. Membranes were incubated in polyclonal rabbit affinity purified HSP70 primary antibody (AS05 061A; Agrisera, Vännäs, Sweden) at a concentration of 1:5000 in 1% milk powder tris-buffered saline with Tween20 (TBS-T) solution for 1 h at room temperature. This antibody is specific to the inducible isoform of salmonid HSP70 and does not detect the constitutive protein, providing a powerful tool to assess induction of HSP70 with thermal challenge. The secondary antibody was goat anti-rabbit immunoglobin G-horseradish peroxidase (IgG-HRP) (SAB 300, Enzo Life Sciences, New York, USA) used at a concentration of 1:5000 in 1% milk powder TBS-T solution for 1 h at room temperature. Chemiluminescent detection of protein bands was performed using ECL Select (GE Healthcare, Buckinghamshire, UK). Blots were imaged using a VersaDoc™ MP 4000 Molecular Imager (Bio-Rad), analysed using Image Lab® software, and expressed relative to the internal standard facilitating qualitative comparisons of constitutive levels and fold-changes among tissues.
Metabolite extractions and assays
Frozen heart and red and white muscle tissues were ground under liquid nitrogen, followed by the addition of perchloric acid (8% PCA with 1 mmol/L EDTA). Samples were vortexed and centrifuged at 5 °C for 3 min at 10 000 rpm, and the supernatant was neutralized to pH 7 with NaOH and centrifuged as before for 1 min. The resulting supernatant was stored at −80 °C until processing. For plasma metabolite extraction, the 8% PCA solution did not contain EDTA. Liver glycogen was assayed as described by Clow et al. (2004), and samples were read on a VERSAmax Tunable Microplate Reader (Molecular Devices Corporation) at 340 nm until absorbance stabilized. Hexokinase (25 μL) was then added to each well, and the absorbance was read after 15–25 min.
Plasma, and heart and white muscle lactate were analysed using an NADH-linked assay in a microplate at 340 nm with a VERSAmax tunable microplate reader, using a glycine buffer commercial assay kit (Sigma-Aldrich Canada Ltd.).
CTmax
We assessed the acute thermal tolerance of Atlantic salmon using an established CTmax protocol, recording the temperature when the fish lost equilibrium after a rapid heat ramp (Becker and Genoway 1979; Fangue et al. 2006; Beitinger and Lutterschmidt 2011). Fish were lightly anaesthetized (0.083 g/L buffered MS-222), weighed (±1 g), measured (±1 mm), and then transferred to experimental boxes (40 cm × 13.5 cm × 11 cm) with a clear Plexiglas® lid. They were left to recover in the experimental blackened PVC boxes for 24 h at 15 °C ± 1 °C before experiments began. Each box had an air stone with water flow at 4 L/min, and the DO remained >85% during all trials. CTmax trials for all control fish were performed at 0900, whereas trials for heat-shocked fish were performed at 0900 or 1300. We found no statistically significant differences between the CTmax values of heat-shocked fish that were done at the different times of day (p = 0.5; two-tailed, unpaired t test).
Atlantic salmon were exposed to either control conditions or a heat shock event as described previously. The control fish (15 °C) were left in the boxes for the same amount of time as the fish exposed to the heat shock. After exposure to the control condition or heat, the temperature was set to increase at a rate of 0.33 °C/min (Becker and Genoway 1979) until the fish lost equilibrium for 2 consecutive seconds. The actual rate of heating in our experimental trials was 0.314 ± 0.006 °C/min. The temperature at which the fish lost equilibrium was recorded, and the temperature was immediately decreased over 20 min to 15 °C.
Statistical analyses
Statistical analyses were performed using SPSS Statistics software (version 19; IBM, Chicago, Illinois, USA). Before analysis, all data were checked for violations of assumptions using the Kolmogorov–Smirnov test for normality and the Levene’s test for homogeneity of variance. If data failed these assumptions, they were transformed using either a square root or logarithmic transformation. We used one-way ANOVAs to determine differences over time in our dependent variables. We used Tukey’s post hoc tests to determine differences among sampling points. We performed a logistical regression analysis (R Studio 3.3.2) to determine possible differences in percent mortality in the CTmax tests. For all analyses, the fiducial limit of significance was set to 0.05. All data are presented as mean ± SEM.
Results
HSP70
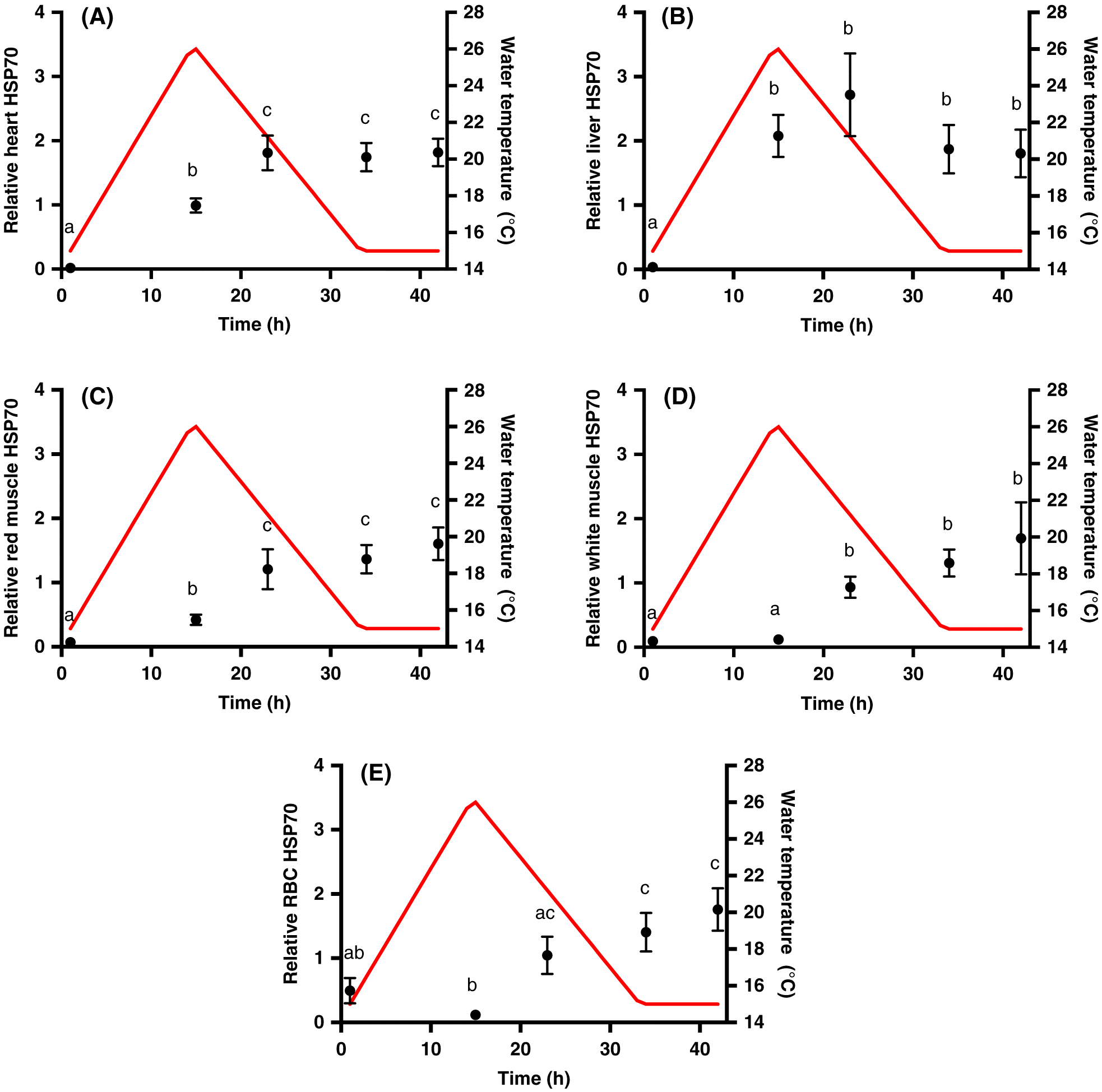
The heat event resulted in a significant increase in inducible HSP70 levels in heart (p < 0.001; Fig. 1A), liver (p = 0.024; Fig. 1B), red muscle (p < 0.001; Fig. 1C), white muscle (p < 0.001; Fig. 1D), and RBCs (p < 0.001; Fig. 1E), with tissue-specific differences in the time of significant induction. There was a significant induction of HSP70 in heart and red muscle at the peak of the thermal cycle (15 h), and a significant increase from the thermal peak to 8 h post peak (23 h). The level of HSP70 then remained constant until 8 h post recovery (42 h). In contrast, we only observed a significant increase in RBC HSP70 from control conditions after 34 h (15 °C recovery). There was also a significant increase in RBC HSP70 from 15–23 h, when the temperature was decreasing back to control levels. Liver HSP70 increased 8 h after the thermal peak (23 h) and returned to control levels in recovery (34 h). In white muscle, HSP70 was significantly induced 8 h following the thermal peak (23 h), and levels remained constant over recovery. The magnitude of the induction of HSP70 was similar across all of the tissues examined.
Metabolic endpoints
There was no significant change in liver glycogen over the course of the heat shock (p = 0.093; Table 1). Glucose and lactate concentrations were measured in both whole blood and plasma to compare results from handheld, whole blood meters and spectrophotometric assays. There were no significant differences between the two methods (glucose p = 0.17; lactate p = 0.21). Thus, we concluded that these portable meters are an acceptable alternative for the measurement of these metabolites in Atlantic salmon. There was a significant transient increase (p < 0.001; Fig. 2A) in plasma glucose concentration at the thermal peak (15 h), which returned to control levels by 8 h after the thermal peak (23 h). We also observed a significant increase in plasma lactate at the thermal peak (15 h), followed by a return to control levels 8 h afterwards (p < 0.001; Fig. 2B). We observed the same trend in heart lactate (p < 0.001; Table 1); however, the concentrations were 5–12× greater in heart tissue than in blood/plasma. Lactate levels in white muscle were significantly elevated from the control level by the peak of the thermal cycle and remained elevated until 8 h after the thermal peak (p = 0.01; Table 1).
Fig. 2.
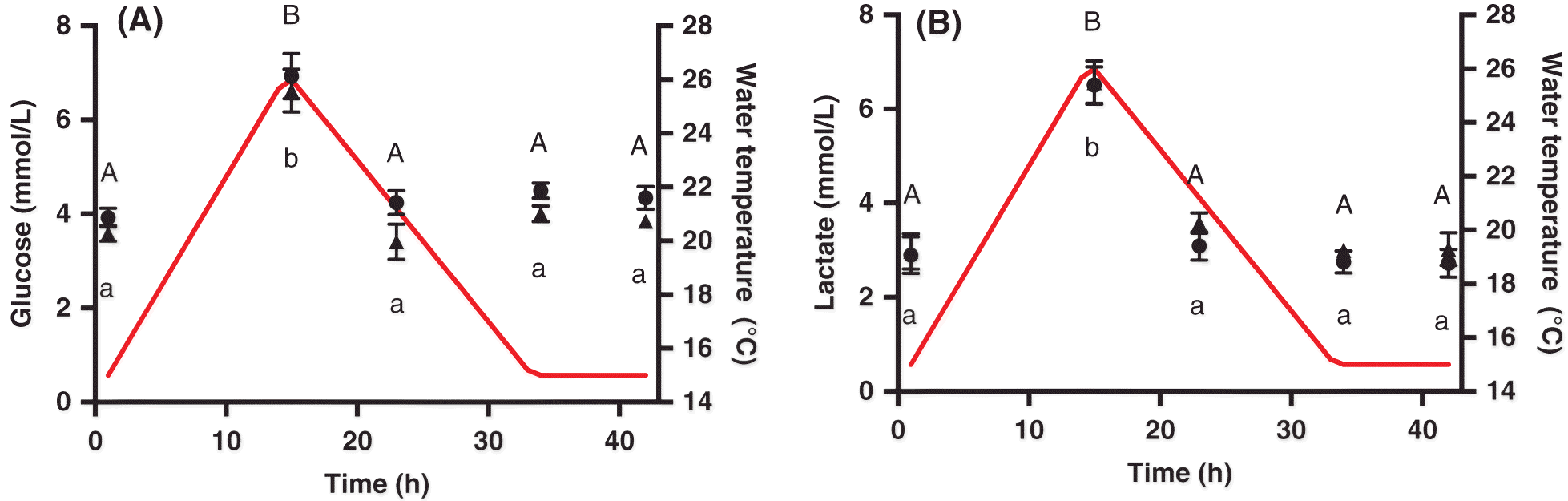
Table 1.
Tissue | End point | Control (t = 0 h) | Peak (t = 15 h) | 8 h post peak (t = 23 h) | Recovery (t = 34 h) | 8 h post recovery (t = 42 h) |
|---|---|---|---|---|---|---|
Liver | Glycogen (μmol glucosyl units/g tissue) | 113.4 ± 55.11a | 106.07 ± 42.52a | 26.48 ± 9.33a | 73.16 ± 20.37a | 39.80 ± 11.05a |
White muscle | Lactate (mmol/L) | 84.19 ± 3.61a | 103.57 ± 4.39b | 112.77 ± 4.29b | 85.74 ± 3.92a | 95.59 ± 4.77ab |
Heart | Lactate (mmol/L) | 32.49 ± 3.30a | 44.84 ± 2.01b | 34.74 ± 2.26a | 38.51 ± 1.52ab | 34.88 ± 3.37a |
Note: Data are presented as mean ± SEM. Values with different lowercase letters indicate significant differences (one-way ANOVA followed by Tukey’s post hoc testing).
CTmax
There was no statistically significant difference between the CTmax of the control (29.3 °C ± 0.55 °C) fish compared with fish exposed to the heat shock (31.0 °C ± 0.4 °C; p = 0.698). However, there was a significant positive correlation between the CTmax of individual fish and their condition factor (p = 0.033; R 2 = 0.3035), in that fish with a higher condition factor tended to have a higher CTmax. There were no differences in the condition factor of the fish used in the control group and the fish experiencing a thermal cycle (p = 0.402). Notably, the mortality rate of the fish that had experienced the heat shock was 55% compared with 33% for the control fish; however, the odds of survival were not statistically different between the two groups.
Discussion
Here, we describe the time course of cellular and metabolic responses in Atlantic salmon to a short-term, acute thermal event representative of field conditions. Our recent work in salmonids has shown distinct physiological and biochemical changes after several days of diel thermal cycling (Callaghan et al. 2016; Tunnah et al. 2017), and we were therefore interested in more detailed stress responses after ∼1 d of a field-relevant heat event. We also assessed whether this heat challenge would affect tolerance to a subsequent heat shock in terms of CTmax. We expected to observe signs of both cellular and physiological stress in these fish, and this prediction was supported. Contrary to our second prediction, we found no significant effect of a prior heat shock on CTmax.
A decrease in energy reserves may also be indicative of a stressed state. Glycogen is an energy storage molecule that can be broken down into glucose in times of high-energy needs, and thus we used this metabolite as an indication of energy status during the heat event. Despite the lack of any obvious decrease in liver glycogen over the heat shock, we did observe signs that the fish were mobilizing energy reserves, as both plasma and whole blood glucose increased. Similarly, white muscle lactate was elevated at the peak of the thermal cycle when metabolic demands should be elevated. Stressed salmon have increased energy demands (Vijayan et al. 1990) and cease feeding at high temperatures (Breau et al. 2011); thus, the temperature increase experienced here would necessitate the mobilization of some energy stores over the 1 d heat shock. Accordingly, fish with a higher condition factor and more energy reserves may be more thermally tolerant, as we observed here with the positive correlation of CTmax with condition factor. Recently, Callaghan et al. (2016) showed that 1 d of sub-critical heat exposure in rainbow trout (Oncorhynchus mykiss) depleted glycogen stores; however, after multiple days of thermal cycling, fish adopted an anabolic phenotype and replenished energy reserves. Such metabolic plasticity would equip Atlantic salmon to cope with subsequent heat exposures and would likely be even more important as temperatures approached their upper limits.
HSP70 induction in brook trout has been correlated with population-specific thermal tolerances (Stitt et al. 2014) and thermal limits (Chadwick et al. 2015), leading to the suggestion that HSPs are useful markers to determine ecological thresholds in salmonids (Chadwick et al. 2015). Notably, two highly stress-inducible HSPs, HSP70 and HSP30, significantly increased in wild Miramichi Atlantic salmon during heat events when water temperatures reached 23 °C (Lund et al. 2002). This cellular stress response coincides with the upper threshold for feeding in Atlantic salmon (Breau et al. 2011) and is likely indicative of protein damage (Tunnah et al. 2017). Fader et al. (1994) showed that HSPs were induced in several species of fishes with natural seasonal variations in temperature and there have been subsequent studies in a variety of species on this theme (e.g., Podrabsky and Somero 2004; Todgham et al. 2006; Narum et al. 2013). Tissue-specific HSP70 induction patterns were observed in Atlantic salmon experiencing natural diel thermal cycles that were distinct from an acute heat shock (Tunnah et al. 2017). Here, we measured induction profiles of HSP70 throughout the heat event, rather than just before and after, in five tissues to understand immediate responses to an ecologically relevant heat stress. The induction was similar in magnitude in all tissues, but we did observe distinct time courses of the heat shock response in each tissue that may reflect tissue-specific sensitivities to thermal stress. For example, the early induction of HSP70 in the heart and red muscle is suggestive of early damage in these highly aerobic tissues.
In this study, HSP70 remained elevated into recovery in all tissues. In brook trout, hsp genes remained elevated at least 48 h following an acute heat shock (Lund et al. 2003), suggesting that recovery from the heat shock response does not occur before the next day of heat cycling. On the one hand, maintaining a high titer of inducible HSPs in tissues will offer protection against thermal denaturation and may precondition tissues against a second thermal insult. Indeed, the induction of HSPs has been correlated with thermal preconditioning in fishes (Feminella and Matthews 1984; Hightower et al. 1999), whereby exposure to a prior heat event protects the animal from a subsequent higher temperature that may otherwise be lethal. However, our CTmax data do not support this possibility, in that fish exposed to ∼1 d thermal cycle were not more thermally tolerant than fish maintained under control conditions. In contrast, given the energetic cost of protein synthesis in cells (∼11–42% of total energy expenditure; Carter and Houlihan 2001), a robust heat shock response represents a significant cost over multiple days/months of warming. Although we found no evidence of enhanced thermal tolerance with 1 d of thermal cycling, several days of warm, ecologically relevant diel cycling has recently been shown to increase CTmax in wild Atlantic salmon (Corey et al. 2017). We note that our mortality rates were high following CTmax trials and this may be linked to the depletion of cardiac glycogen stores after a single thermal cycle, as we observed previously (Callaghan et al. 2016), and (or) irreversible cell and tissue damage. These factors could increase the sensitivity of the heart to arrhythmias at the CTmax temperature (Anttila et al. 2013). Although Atlantic salmon demonstrated cardiac plasticity with temperature (Anttila et al. 2014), their capacity for physiological plasticity in their upper thermal limits may be small relative to their lower thermal limits, as observed in other fish (Sandblom et al. 2016).
In July 2010, the water temperature of the Miramichi River remained above 23 °C, the critical temperature of Atlantic salmon, for 18 consecutive days (DFO 2012). High mortality and the eventual closing of the recreational fishery followed this temperature event. Here, we show that a single thermal event, representative of summer water temperatures in the Miramichi River, resulted in heat shock and physiological stress responses, and increased energy demands with no effect on acute thermal tolerance. Atlantic salmon can cope with this short-term heat shock, but this challenge may compromise the ability to cope with a later acute heat stress. This conclusion is supported by the lack of any obvious thermal preconditioning in wild Atlantic salmon experiencing several days of thermal cycling and then subjected to an acute heat shock (Tunnah et al. 2017). Recognizing the thermal limits, tolerance, and physiological consequences of thermal stresses faced by Atlantic salmon is critical to manage this important species and to understand the biological impacts of recorded and anticipated increases in temperature.
Acknowledgements
This study was supported by NSERC Discovery Grants (SC, TJM) and the Harold Crabtree Foundation (SC). The authors would like to enthusiastically thank Wayne Anderson for animal husbandry, David J. Summerby-Murray and Luke Poirier for analytical assistance, Dr. Cindy Breau Fisheries and Oceans (DFO) for helpful discussions, and the Fisheries and Oceans Mactaquac Hatchery, NB, Canada for supplying fish.
References
Anttila K, Casselman MT, Schulte PM, and Farrell AP. 2013. Optimum temperature in juvenile salmonids: connecting subcellular indicators to tissue function and whole-organism thermal optimum. Physiological and Biochemical Zoology, 86: 245–256.
Anttila K, Couturier C, Overli O, Johnson A, Marthinsen G, Nilsson GE, et al. 2014. Atlantic salmon show capability for cardiac acclimation to warm temperatures. Nature Communications, 5: 4252.
Becker CD, and Genoway RG. 1979. Evaluation of the critical thermal maximum for determining thermal tolerance in freshwater fish. Environmental Biology of Fishes, 4: 245–256.
Beitinger TL, and Lutterschmidt WI. 2011. Measures of thermal tolerance. In Encyclopedia of fish physiology: from genome to environment. Edited by AP Farrell. Academic Press, Elsevier Inc., London, UK. pp. 1695–1702.
Breau C, Cunjak RA, and Bremset GG. 2007. Age-specific aggregation of wild juvenile Atlantic salmon (Salmo salar) at cool water sources during high temperature events. Journal of Fish Biology, 71: 1179–1191.
Breau C, Cunjak RA, and Peake SJ. 2011. Behaviour at high water temperatures: can physiology explain movement of juvenile Atlantic salmon (Salmo salar) to cool water? Journal of Animal Ecology, 80: 844–853.
Caissie D, Breau C, Hayward J, and Cameron P. 2012. Water temperature characteristics within the Miramichi and Restigouche rivers. Department of Fisheries and Oceans Canadian Science Advisory Secretariat, Academic Press, Canada. pp. 1–38.
Caissie D, El-Jabi N, and Turkkan N. 2014. Stream water temperature modeling under climate change scenarios B1 & A2. Department of Fisheries and Oceans Canadian Technical Report of Fisheries and Aquatic Sciences 3106, Canada.
Callaghan N, Tunnah L, Currie S, and MacCormack TJ. 2016. Metabolic adjustments to short-term diurnal temperature fluctuation in the rainbow trout (Oncorhynchus mykiss). Physiological and Biochemical Zoology, 89(6): 498–510.
Carter CG, and Houlihan DF. 2001. Protein synthesis. In Fish physiology: nitrogen excretion. Edited by PA Wright, and PM Anderson. Academic Press, New York, New York. pp. 31–75.
Chadwick JG, Nislow KH, and McCormick SD. 2015. Thermal onset of cellular and endocrine stress response correspond to ecological limits in brook trout, an iconic cold-water fish. Conservation Physiology, 3(1): cov017.
Clow A, Thorn L, Evans P, and Hucklebridge F. 2004. The awakening cortisol response: methodological issues and significance. Stress, 7(1): 29–37.
Corey E, Linnansaari T, Cunjak RA, and Currie S. 2017. Physiological effects of environmentally relevant, multi-day thermal stress on wild juvenile Atlantic salmon (Salmo salar). Conservation Physiology, 5(1): cox01410.
COSEWIC. 2010. Atlantic salmon (Salmo salar): inner Bay of Fundy population [online]: Available from sararegistry.gc.ca/virtual_sara/files/cosewic/sr_atlantic_salmon_inner_bay_e.pdf.
Currie S, and Schulte PM. 2014. Thermal stress. In The physiology of fishes. 4th edition. Edited by DH Evans, JB Claiborne, and S Currie. CRC Press, Boca Raton, Florida. pp. 257–287.
Currie S, Moyes CD, and Tufts BL. 2000. The effects of heat shock and acclimation temperature on hsp70 and hsp30 mRNA expression in rainbow trout: in vivo and in vitro comparisons. Journal of Fish Biology, 56(2): 398–408.
Department of Fisheries and Oceans (DFO). 2012. Temperature threshold to define management strategies for Atlantic salmon (Salmo salar) fisheries under environmentally stressful conditions. DFO Canadian Science Advisory Report, 2012: 1–18.
Department of Fisheries and Oceans (DFO). 2013. Atlantic salmon (Salmo salar) returns to the Miramichi River (NB) for 2012. DFO Canadian Science Advisory Secretariat Science Response, 2013/009.
DuBeau SF, Pan F, Tremblay GC, and Bradley TM. 1998. Thermal shock of salmon in vivo induces the heat shock protein hsp70 and confers protection against osmotic shock. Aquaculture, 168(1): 311–323.
Eldridge WH, Sweeney BW, and Law JM. 2015. Fish growth, physiological stress, and tissue condition in response to rate of temperature change during cool or warm diel thermal cycles. Canadian Journal of Fisheries and Aquatic Sciences, 72: 1527–1537.
Fader SC, Yu Z, and Spotila JR. 1994. Seasonal variation in heat shock proteins (hsp 70) in stream fish under natural conditions. Journal of Thermal Biology, 19(5): 335–341.
Fangue NA, Hofmeister M, and Schulte PM. 2006. Intraspecific variation in thermal tolerance and heat shock protein gene expression in common killifish, Fundulus heteroclitus. Journal of Experimental Biology, 209: 2859–2872.
Feminella JW, and Matthews WJ. 1984. Intraspecific differences in thermal tolerance of Etheostoma spectabile (Agassiz) in constant versus fluctuating environments. Journal of Fish Biology, 25: 455–461.
Fowler SL, Hamilton D, and Currie S. 2009. A comparison of the heat shock response in juvenile and adult trout (Oncorhynchus mykiss)—implications for increased thermal sensitivity with age. Canadian Journal of Fisheries and Aquatic Sciences, 66: 91–100.
Fry F. 1958. Temperature compensation. Annual Review of Physiology, 20: 207–224.
Galloway BJ, and Kieffer JD. 2003. The effects of an acute temperature change on the metabolic recovery from exhaustive exercise in juvenile Atlantic salmon (Salmo salar). Physiological and Biochemical Zoology, 76(5): 652–662.
Hightower LE, Norris CE, DiIorio PJ, and Fielding E. 1999. Heat shock responses of closely related species of tropical and desert fish. American Zoology, 39: 877–888.
Hochachka PW, and Somero GN. 2002. Biochemical adaptation: mechanism and process in physiological evolution. Oxford University Press, New York, New York.
Houston AH, and Gingras-Bedard JH. 1994. Variable versus constant temperature acclimation regimes: effects on hemoglobin isomorph profile in goldfish, Carassius auratus. Fish Physiology and Biochemistry, 13(6): 445–450.
Kolhatkar A, Robertson CE, Thistle ME, Gamperl AK, and Currie S. 2014. Coordination of chemical (trimethylamine oxide) and molecular (heat shock protein 70) chaperone responses to heat stress in elasmobranch red blood cells. Physiological and Biochemical Zoology, 87(5): 652–662.
LeBlanc S, Middleton S, Gilmour KM, and Currie S. 2011. Chronic social stress impairs thermal tolerance in the rainbow trout (Oncorhynchus mykiss). Journal of Experiment Biology, 214: 1721–1731.
Lund SG, Caissie D, Cunjak RA, Vijayan MM, and Tufts BL. 2002. The effects of environmental heat stress on heat shock mRNA and protein expression in Miramichi Atlantic salmon (Salmo salar) parr. Canadian Journal of Fisheries and Aquatic Sciences, 59: 1553–1562.
Lund SG, Lund MEA, and Tufts BL. 2003. Red blood cell Hsp70 mRNA and protein as bioindicators of temperature stress in the brook trout (Salvelinus fontinalis). Canadian Journal of Fisheries and Aquatic Sciences, 60: 460–470.
Mesa M, Weiland L, and Wagner P. 2002. Effects of acute thermal stress on the survival, predator avoidance, and physiology of juvenile fall chinook salmon. Northwest Science, 76: 118–128.
Narum SR, Campbell NR, Meyer KA, Miller MR, and Hardy RW. 2013. Thermal adaptation and acclimation of ectotherms from differing aquatic climates. Molecular Ecology, 22(11): 3090–3097.
Podrabsky JE, and Somero GN. 2004. Changes in gene expression associated with acclimation to constant temperatures and fluctuating daily temperatures in an annual killifish Austrofundulus limnaeus. Journal of Experimental Biology, 207: 2237–2254.
Sandblom E, Clark TD, Gräns A, Ekstrom A, Brijs J, Sundström F, et al. 2016. Physiological constraints to climate warming in fish follow principles of plastic floors and concrete ceilings. Nature Communications, 7: 11447.
Stevens ED, and Sutterlin AM. 1976. Heat transfer between fish and ambient water. Journal of Experimental Biology, 65(1): 131–145.
Stitt BC, Burness G, Burgomaster KL, Currie S, McDermid JL, and Wilson CC. 2014. Intraspecific variation in thermal tolerance and acclimation capacity in brook trout (Salvelinus fontinalis): physiological implications for climate change. Physiological and Biochemical Zoology, 87: 15–29.
Threader RW, and Houston AH. 1983. Heat tolerance and resistance in juvenile rainbow trout acclimated to diurnally cycling temperatures. Comparative Biochemistry and Physiology Part A: Physiology, 75(2): 153–155.
Todgham AE, Iwama GK, and Schulte PM. 2006. Effects of the natural tidal cycle and artificial temperature cycling on Hsp levels in the tidepool sculpin Oligocottus maculosus. Physiological and Biochemical Zoology, 79: 1033–1045.
Tunnah L, Currie S, and MacCormack TJ. 2017. Do prior diel thermal cycles influence the physiological response of Atlantic salmon (Salmo salar) to subsequent heat stress? Canadian Journal of Fisheries and Aquatic Sciences, 74(1): 127–139.
Turnbull J, Bell A, Adams C, Bron J, and Huntingford F. 2005. Stocking density and welfare of cage farmed Atlantic salmon: application of a multivariate analysis. Aquaculture, 243: 121–132.
Vijayan MM, Ballantyne JS, and Leatherland JF. 1990. High stocking density alters the energy metabolism of brook charr, Salvelinus fontinalis. Aquaculture, 88(3): 371–381.
Information & Authors
Information
Published In

FACETS
Volume 2 • Number 1 • January 2017
Pages: 330 - 341
Editor: Karen Kidd
History
Received: 23 September 2016
Accepted: 6 February 2017
Version of record online: 4 April 2017
Copyright
© 2017 Gallant et al. This work is licensed under a Creative Commons Attribution 4.0 International License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Data Availability Statement
All relevant data are within the paper.
Key Words
Sections
Subjects
Plain Language Summary
Beat the heat: Physiological responses of Atlantic salmon to an ecologically relevant thermal challenge
Authors
Author Contributions
MJG and SC conceived and designed the study.
MJG performed the experiments/collected the data.
All analyzed and interpreted the data.
All contributed resources.
All drafted or revised the manuscript.
Competing Interests
The authors have declared that no competing interests exist.
Metrics & Citations
Metrics
Other Metrics
Citations
Cite As
Melanie J. Gallant, Sacha LeBlanc, Tyson J. MacCormack, and Suzanne Currie. 2017. Physiological responses to a short-term, environmentally realistic, acute heat stress in Atlantic salmon, Salmo salar. FACETS.
2: 330-341.
https://doi.org/10.1139/facets-2016-0053
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
Cited by
1. Differences in gene expression between high and low tolerance rainbow trout (Oncorhynchus mykiss) to acute thermal stress
2. Wild juvenile Atlantic salmon (Salmo salar) offer insights into movement patterns of territorial freshwater fishes in relation to high temperature and proximity to thermal refuges
3. A case study on the effect of aquaculture operations on the physiology and behaviour of Atlantic salmon (Salmo salar) during two heat events on a commercial farm
4. Characterisation of Fatty Acid Profiles in Tasmanian Atlantic Salmon Muscle, Oocytes and Eggs in Differing Fully Commercial Settings
5. Long-term substitution of fish oil with alternative sources in Atlantic salmon (Salmo salar): Performance, health, and consumer appeal
6. Effect of increasing ambient temperature on physiological changes, oxidative stress, nitric oxide, total antioxidant power, and mitochondrial activity of Nile tilapia (Oreochromis niloticus Linn.)
7. Non‐concordant epigenetic and transcriptional responses to acute thermal stress in western mosquitofish (
Gambusia affinis
)
8. Impacts of Climate Change on the Parasite–Host Relationship in Freshwater Aquaculture
9. The impacts of diel thermal variability on growth, development and performance of wild Atlantic salmon (
Salmo salar
) from two thermally distinct rivers
10. Considerations for Water Temperature-Related Fishery Closures in Recreational Atlantic Salmon (
Salmo salar
) Catch and Release Fisheries: A Case Study from Eastern Canada
11. Characterising the Physiological Responses of Chinook Salmon (Oncorhynchus tshawytscha) Subjected to Heat and Oxygen Stress
12. The upper temperature and hypoxia limits of Atlantic salmon (
Salmo salar
) depend greatly on the method utilized
13. The Atlantic salmon's (Salmo salar) incremental thermal maximum is a more relevant and sensitive indicator of family-based differences in upper temperature tolerance than its critical thermal maximum
14. The Role of Cold-Water Thermal Refuges for Stream Salmonids in a Changing Climate—Experiences from Atlantic Canada
15. Combined effects of rearing temperature regime (thermocycle vs. constant temperature) during early development and thermal treatment on Nile tilapia (Oreochromis niloticus) sex differentiation
16. Effects of juvenile thermal preconditioning on the heat-shock, immune, and stress responses of rainbow trout upon a secondary thermal challenge
17. Lowering temperature to 1 °C results in physiological changes, stress and mortality in cultured Atlantic Salmon (Salmo salar)
18. Combined effects of high temperature and pesticide mixture exposure on free-swimming behaviors and hepatic cytochrome P450 1A expression in goldfish,
Carassius auratus
19. Short-term acclimation dynamics in a coldwater fish
20. Change in plasma cortisol and glucose levels of Oncorhynchus
keta according to water temperature
21. Timing and frequency of high temperature events bend the onset of behavioural thermoregulation in Atlantic salmon (
Salmo salar
)
22. Differential heat shock protein responses in two species of Pacific salmon and their utility in identifying heat stress
23. Increased Thermal Challenges Differentially Modulate Neural Plasticity and Stress Responses in Post-Smolt Atlantic Salmon (Salmo salar)
24. Effects of heat shock on energy metabolism and antioxidant defence in a tropical fish species
Psalidodon bifasciatus
25. Acute critical thermal maximum does not predict chronic incremental thermal maximum in Atlantic salmon (Salmo salar)
26. Understanding risks and consequences of pathogen infections on the physiological performance of outmigrating Chinook salmon
27. The Atlantic salmon’s stress- and immune-related transcriptional responses to moderate hypoxia, an incremental temperature increase, and these challenges combined
28. A bright spot analysis of inland recreational fisheries in the face of climate change: learning about adaptation from small successes
29. Temperature increase and its effects on fish stress physiology in the context of global warming
30. The physiological ups and downs of thermal variability in temperate freshwater ecosystems
31. Rearing temperature conditions (constant vs. thermocycle) affect daily rhythms of thermal tolerance and sensing in zebrafish
32. Comparative study on the non-specific immune response and hsp70 gene expression among three strains of koi (Cyprinus carpio) under acute heat stress
33. Mimicking Transgenerational Signals of Future Stress: Thermal Tolerance of Juvenile Chinook Salmon Is More Sensitive to Elevated Rearing Temperature Than Exogenously Increased Egg Cortisol
34. Upper thermal limits are repeatable in Trinidadian guppies
35. Thermal injuries in Atlantic salmon in a pilot laboratory trial
36. Acute exposure of larval rainbow trout (Oncorhynchus mykiss) to elevated temperature limits hsp70b expression and influences future thermotolerance
37. Multiple sub-lethal thresholds for cellular responses to thermal stressors in an estuarine fish
38. Acclimation capacity of the cardiac HSP70 and HSP90 response to thermal stress in lake trout (Salvelinus namaycush), a stenothermal ice-age relict
39. Heat tolerance and gene expression responses to heat stress in threespine sticklebacks from ecologically divergent environments