Bat bites and rabies: the Canadian scene
Abstract
Bats are susceptible to rabies. Although bats may appear to be asymptomatic carriers of rabies for a few days, eventually they fall ill to the viral infection and die. Two of at least four bat-specific variants of rabies virus in Canada have killed humans. Rabies is usually transmitted by biting, but bats are small mammals so their bites may go unnoticed. People exposed to rabid animals should receive postexposure prophylaxis (PEP). With 60 known human deaths from 1950 to 2009, rabies is rare in Canada and the United States of America compared with India where it kills over 100 people annually. In Asia and Africa, most human rabies is acquired from dog bites. In Brazil, dog and bat bites together account for >80% of human rabies. In Canada, rabies is a disease primarily confined to wildlife (foxes, racoons, skunks, and bats). The public image of bats is negatively affected by their association with diseases. Too often bats are victimized by allegations of their role in deadly diseases such as rabies, Ebola, severe acute respiratory syndrome (SARS), and Middle East respiratory syndrome (MERS). In general, bats are not dangerous, but humans should seek treatment if they are bitten by one. (Graphical abstract shows a 4-g elegant myotis biting MBF’s finger—photo by Sherri and Brock Fenton.)
Graphical Abstract
Introduction
Rabies, an acute disease of the nervous system, is caused by a Lyssavirus (Fooks and Jackson 2020). Worldwide, rabies kills about 59 000 people a year, with 95% of human deaths occurring in Asia and Africa (Hampson et al. 2015). Typically, the rabies virus is spread by dog bites; at the bite wound the victim is inoculated with infectious virus in the dog’s saliva. The tally of human deaths from rabies varies dramatically across countries. For example, in India from 2012 to 2014 there were 128 (42.7 per year; Mani et al. 2016), in South Africa from 2008 to 2018 there were 104 (10.4 per year; Dermaux-Msimang et al. 2019), and in Brazil from 2000 to 2017 there were 188 human rabies cases (27 per year; Vargas et al. 2019). What about the association of bats and rabies?
Rabies is also spread by bats (Fig. 1). In Brazil, rabies from bats accounted for >40% of the 188 human cases and from dogs about another 40% (Vargas et al. 2019). Bat-related rabies in Brazil is contracted from either blood-feeding vampire bats (Fig. 1C) or insectivorous bats. Vampire bat bites on humans usually result from blood feeding, whereas bites from insectivorous bats (e.g., Brazilian free-tailed bat, Tadarida brasiliensis) are normally delivered in self-defense but also can be from aggressive rabid bats. Escobar et al. (2015) noted a considerable reduction in rabies in dogs in Brazil, while the numbers of cases of bat rabies has remained steady for about 10 years. They noted that bats appeared to be the principal reservoir for rabies in the Neotropics.
Fig. 1.

Rabies in humans is much less common in the USA and Canada where, from 1950 to 2009, there were 60 cases of rabies acquired domestically (Fenton et al. In press). Bat rabies variants caused a number of these fatalities, but 33% involved no history of exposure to bats, and in 28% there was no report of direct contact with a bat (De Serres et al. 2008; Jackson 2011a). In Canada, seven of the nine most recent human rabies fatalities were caused by bat virus variants (Table 1). Notably, none of these involved rabies virus variants associated with big brown bats (Fig. 1A), which is perhaps the most common house bat across southern Canada. Details of two recent human deaths from bat rabies illustrate the complexity of the situation.
Table 1.
| Year | Patient age/sex | Province | Vector/virus variant | References |
|---|---|---|---|---|
| 1970 | 15/male | Saskatchewan | Bat/? | Dempster et al. 1972 |
| 1977 | 63/male | Nova Scotia | Bat/? | King et al. 1978 |
| 1984 | 43/male | Quebec | Dog (Dominican Republic)/? | Picard 1984; Webster et al. 1985 |
| 1985 | 22/male | Alberta | Bat/? | Dolman and Charlton 1987; Webster et al. 1987 |
| 2000 | 9/male | Quebec | Bat/variant associated with silver-haired bat | Turgeon et al. 2001; Despond et al. 2002; Elmgren et al. 2002 |
| 2003 | 52/male | British Columbia | Bat/variant associated with Myotis spp. bat | Parker et al. 2003; Walker et al. 2016 |
| 2007 | 73/male | Alberta | Bat/variant associated with silver-haired bat | Johnstone et al. 2008; McDermid et al. 2008 |
| 2012 | 41/male | Ontario | Dog (Dominican Republic)/variant associated with Haitian dog | Dyer et al. 2013; Wilcox et al. 2014 |
| 2019 | 21/male | British Columbia | Bat/variant associated with silver-haired bat | Brown 2019; K. Knowles, 2019, personal communication, Canadian Food Inspection Agency, Fallowfield, Ontario, Canada |
Note: Shown are the year of fatality, age and sex of the patient, province where the infection occurred, disease transmitting organism (vector) and strain (variant) of rabies virus isolated from patient, if known. Adapted with permission from Jackson (2011b).
In Montreal in 2000, a 9-year-old boy died of rabies caused by the silver-haired bat (L. noctivagans, Fig. 1B) virus variant. This variant also is known from the tricoloured bat (Perimyotis subflavus) (Turgeon et al. 2001; Despond et al. 2002; Elmgren et al. 2002). This case illustrates four issues presented by bats and rabies. First, the two bat species associated with the silver-haired rabies virus variant rarely roost in buildings and thus are expected to have infrequent human contact. Second, the boy and his family had stayed in a rented cottage in a park in Quebec. Two bats, identified by the parents as little brown myotis (Myotis lucifugus), were captured in the cottage by the parents and released outside. Third, the victim had a slight abrasion on his upper arm, which in retrospect may have been a bat bite, but apart from this scrape there was no report or clear evidence of a bite. Two of us (MBF and ACJ) visited the cottage in December 2000, and careful searching revealed no physical evidence (e.g., droppings) that bats had roosted in the cottage. But another cottage within 1 km of the site was used as a roost by bats, based on the accumulation of droppings under specific roosting sites. Fourth, did the bat exposure happen in the cabin, and if so, how could bats have entered the cottage occupied by the boy and his family? Could bats possibly have come into the house on firewood or flown down the metal chimney into the wood stove? Unfortunately, the lack of details precludes drawing definitive conclusions about how the boy’s exposure to a rabid bat had occurred.
In 2019 in British Columbia, a 21-year-old man died of rabies caused by the silver-haired bat virus variant. This was the first human death in Canada from indigenously acquired rabies since 2007 (Table 1). The victim had been exposed to a bat on Vancouver Island around the middle of May and showed clinical symptoms of rabies 6–8 weeks afterwards. The exposure route is unknown, possibly from a scratch rather than a bite. The time lag between exposure and appearance of symptoms is typical.
Recognizing a bat exposure
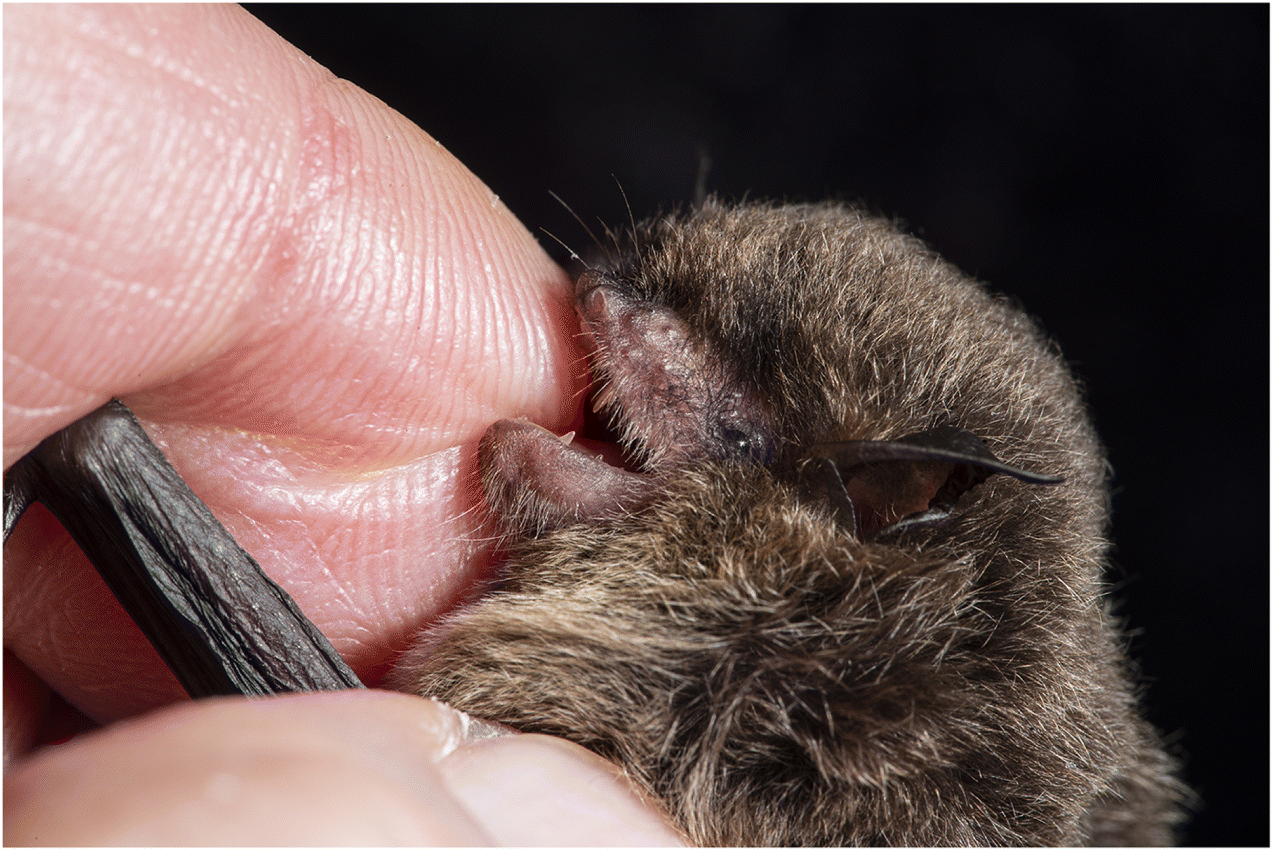
Normally, a bite from a wild mammal indicates possible exposure to a rabid animal. But bat bites can be hard to recognize (Fig. 2). When do bats bite? Normally bats, like most mammals, bite only in self-defense. Even the three species of vampire bats that bite to eat (drink) blood also bite in self-defense. Bats expressing clinical signs of furious rabies may bite indiscriminately and without provocation (see below). These bites and defensive bites usually leave puncture wounds, which may be small and difficult to recognize when inflicted by an insectivorous bat (e.g., Figs. 2A, 2C). The blade-like upper incisor and canine teeth of vampire bats slice rather than puncture tissue and leave obvious, bleeding wounds (Fig. 2B).
Fig. 2.

Bats are among the smallest mammals that are rabies vectors (Fenton and Simmons 2015). This has important public health implications because their small size can make bat bites hard to notice. For example, it is easy to overlook the bite inflicted by an adult silver-haired bat with a mass of about 9.5 g (Jackson and Fenton 2001), which could place the victim in jeopardy (Table 1). About 20 bat species have been reported in Canada (Van Zyll de Jong 1985), with hoary bats (Lasiurus cinereus; ∼30 g) being the largest and eastern small-footed bats (Myotis leibii; ∼4 g) among the smallest. Hoary bat bites are obvious and painful, while those from eastern small-footed bats may not break the skin (e.g., Fig. 2C).
Any mammal, including bats, will bite while in the grip of furious rabies. Any unprovoked bat bite should be treated as a potential exposure to rabies. The best way to confirm the diagnosis of rabies is by submitting the head of the animal (in the case of bats, the whole body) to a certified laboratory for detection of rabies virus antigen in the brain. This also facilitates accurate identification of the species involved. But if the animal that delivered the bite is not available, how is it possible to confirm exposure especially if there is no evidence of a bite?
Diagnostic laboratory tests for rabies in humans or animals include detection of rabies virus antigen in post-mortem brain tissue. In humans, antemortem diagnostic tests include detection of rabies virus antigen in skin biopsies or rabies virus RNA using the reverse transcription polymerase chain reaction in saliva, skin biopsies obtained from the nape of the neck, and cerebrospinal fluid (Jackson 2020a). The presence of serum neutralizing anti-rabies virus antibodies is also of diagnostic value in previously unvaccinated individuals (also in cerebrospinal fluid whether previously vaccinated or not).
Humans directly exposed to a known or suspected rabid animal should receive rabies post-exposure prophylaxis (PEP) treatment. Exposures to bat rabies could occur through bites, scratches, or having infected saliva contact abrasions or cuts in the skin (NACI 1998). Since the mid-1990s, PEP has also been administered to persons sleeping unattended in a room with a bat when the individual was unable to exclude the possibility of a bat “exposure” and the bat in question was unavailable for testing. Over time, this recommendation had been expanded to include additional situations where persons are unable to report possible bat exposures, such as children or intoxicated/cognitively impaired individuals in the same room as a bat (NACI 2002).
The cost and questionable benefits of widespread administration of rabies PEP (Huot et al. 2008; De Serres et al. 2009) led the National Advisory Committee on Immunization to recommend PEP only when there has been direct human contact with a bat that is unavailable for testing or when a possible bat exposure (e.g., bite; scratch; saliva from a bat contacting a wound or skin abrasion; or unattended sleeping person, child or intoxicated/cognitively impaired individual in the same room as a bat) cannot be excluded (NACI 2009; Warshawsky and Desai 2010).
Could these measures have saved either victim? We think not. Even though neither person reported directly handling or intentionally touching a bat, both victims (or their families) later recalled the victim having some type of bat incident or possible exposure. Almost all humans who have died from rabies in Canada and elsewhere did not receive PEP before the onset of clinical symptoms of rabies.
Rabies kills bats
To the best of our knowledge, all bats are susceptible to rabies and once a bat shows clinical signs of rabies, the illness is fatal (Rupprecht et al. 2002). Nadin-Davis et al. (2001) molecularly characterized rabies virus variants from bat species native to Canada and found four principal phylogenetic groups. Group I included colonial species such as mouse-eared bats (Myotis spp.) and the big brown bat. Group II included the so-called migratory bats: the eastern red bat (Lasiurus borealis), hoary bat, and silver-haired bat. Group III variants circulate in big brown bats and likely emerged more recently in this host. Group IV includes some insectivorous and hematophagous bats of Latin America.
Nadin-Davis et al. (2001) found remarkable levels of spillover of rabies virus variants into mouse-eared bats (genus Myotis) from bats of other genera. The silver-haired bat variant showed the greatest level of interspecies spillover (6/19 or 32%) (Nadin-Davis et al. 2001). Spillover of bat rabies virus variants into other mammal species are less common. In Canada, mammals such as cats (Felis catus), foxes (Vulpes fulva), raccoons (Procyon lotor), and skunks (Mephitis mephitis) sometimes catch and eat bats, may be bitten in the process, and may contract rabies if the bat was rabid. In Brazil there are records of cats becoming rabid from interactions with bats (saude.gov.br/o-ministro/961-saude-de-a-a-z/raiva/41858-situacao-epidemiologica).
We lack comprehensive data about the natural incidence of rabies in bats in Canada. Most data on rabies surveillance is passive. The most accurate (and exceptional) data to date come from bats killed at wind turbines that later tested positive for rabies; Klug et al. (2011) found a low incidence of rabies in silver-haired (1%, n = 96) and hoary bats (0%, n = 121). The most commonly reported data on the incidence of rabies comes from bats submitted for rabies testing, which is arguably not a random or representative sample from natural bat populations. We suspect that the proportion of bats naturally infected with rabies virus is quite low and probably has changed little over time even though rabies surveillance and the absolute number of animals submitted for testing has increased (Constantine 1967; Baer and Adams 1970; Bradley 1979; Rosatte 1987).
Signs of rabies
The rabies virus infects the central nervous system, causing mild inflammation of the brain (encephalitis) and spinal cord (myelitis) without prominent degeneration affecting neuronal cell bodies (Rossiter and Jackson 2020). Rabies causes a marked degeneration of neuronal processes, including axons and dendrites due to oxidative stress caused by mitochondrial dysfunction (Jackson 2020b). The onset of rabies in bats is characterized by a short (1–3 d) prodromal phase of nonspecific signs, which then progresses to severe neurologic disease before the onset of coma, respiratory failure, and death (Courter 1954). Rabies is manifested in two distinct clinical forms—furious (encephalitic) rabies and dumb (paralytic) rabies—and the underlying pathogenesis responsible for each remains poorly understood (Jackson 2020a). There are case reports of furiously rabid bats attacking and biting humans (e.g., Constantine 1967; Brass 1994), and at least one of us (PAF) has seen big brown bats with furious rabies bite aggressively and indiscriminately.
A bat expressing furious rabies is easier for lay persons to recognize because it is likely to be hyperactive, aggressive, irritable, or restless and fly erratically. It may make unproved attacks on other animals, including humans or inanimate objects (e.g., its cage). Furiously rabid bats are agitated and very sensitive to sensory stimuli. They are prone to excessive biting, including self-mutilation and hang-on biting, which likely increases the probability of the bite wound being inoculated with infectious saliva and thus successful disease (virus) transmission. Furious rabies occurs in solitary and colonial bats, although it may be more common in solitary or tree-roosting species (Bell 1980; Schowalter 1980; Kuzmin and Rupprecht 2007; Constantine 2009). Furious rabies in bats is more often observed after intracranial laboratory inoculations of rabies virus compared to bats infected intramuscularly, intradermally, or naturally from other infected bats (Baer and Bales 1967; Constantine 1967).
Bats expressing dumb (paralytic) rabies often show moderate to severe paralysis, losing motor coordination and becoming ataxic with tremors and convulsions. These signs could reflect a greater burden of involvement in the peripheral motor nerves, nerve roots, and spinal cord in paralytic rabies than in encephalitic rabies (Jackson 2020a). Bats with dumb rabies may appear abnormally tame, friendly, inquisitive, and (or) placid. These behavioural changes can cause the sick bat to approach conspecifics or other animals (including humans) without fear (and vice versa), thereby placing the rabid bat closer to another animals, which facilitates biting and disease transmission. Paralytic rabies may be more common in highly social colonial species (Constantine et al. 1968; Kuzmin and Rupprecht 2007). Arguably dumb rabies is more dangerous to people than furious rabies: it is easy for a sympathetic and unsuspecting human to recognize an animal that is in distress, paralyzed and (or) otherwise helpless, and try to come to its aid and get bitten in the process.
Rabies and bats
Not all bats exposed to rabies virus develop disease because bats can have lethal or nonlethal infections. After an exposure to rabies virus, bats may not have clinical disease because they develop neutralizing serum anti-rabies virus antibodies that confer protective immunity and result in an abortive infection (Franka et al. 2008). Maternally derived serum-neutralizing anti-rabies virus antibodies cross the placenta and presumably confer temporary protective immunity to newborn young (Constantine et al. 1968).
The incubation period for rabies virus in insectivorous bats is highly variable. Longer incubation periods may facilitate higher viral titres in the salivary glands of infected animals and thus disease transmission (Baer and Bales 1967). Some bats shed virus in their saliva for 12–15 d before showing clinical signs (Constantine 1971). Bats inoculated experimentally in the laboratory with “fixed” rabies (laboratory-adapted) virus strains tend to exhibit shorter incubation times than bats infected with “street virus” (wild type) from natural sources. This could reflect use of higher than natural viral doses in the lab as well as rapid uptake and direct spread in peripheral nerves. Experimentally inoculated bats can have incubation times from 5 to 14 d and even 6 months (Baer and Bales 1967). Incubation periods in bats captured from the wild can be longer (e.g., 28–209 d—Moore and Raymond 1970; Shankar et al. 2004). Laboratory infection studies reveal that viral replication slows and incubation times are longer in cooler temperatures (Sadler and Enright 1959; Sulkin et al. 1960), suggesting that the onset of disease can be prolonged by hibernation. This could produce seasonal variation in the cycle of infectivity which, in turn, may facilitate long term maintenance of rabies virus in some bat populations (Constantine 2009; George et al. 2011).
Recent studies using modern methods for detecting rabies virus infection indicate that bats with clinical signs of rabies do not recover from disease. This contradicts earlier suggestions that bats were asymptomatic carriers of rabies because they could transmit infectious virus but show no clinical signs of illness and not succumb to the disease. Inoculated bats that remain healthy and have survived infection are not carriers of rabies (Moreno and Baer 1980; Franka et al. 2008). Moreover, bats and other mammals can shed infectious virus in their saliva several days (or even weeks) before they begin to show signs of disease onset. But once neurological signs of rabies are evident, the outcome is an inevitable progression to death.
How do bats get rabies? In most cases, probably from bites by other bats (e.g., Bell 1980). The tendency of many bat species to roost together and form social colonies may make them especially vulnerable to rabies transmission. In summer, bats that roost with others (usually conspecifics) often do so to keep warm (Willis and Brigham 2007). Big brown bats and other colonial species live in fission–fusion societies in which individuals roost with an extended group of roost-mates. On any given day, the individuals comprising a “social group” within a roost may differ from the composition of the day before or the group composition one week later. Individuals have a repertoire of roosts and roost mates, potentially facilitating transfer of rabies virus among individual conspecifics and, possibly, heterospecifics (Nadin-Davis et al. 2010). Recent work with proximity tags reveals longer term associations between individuals (e.g., mother and young; Ripperger et al. 2019), perhaps altering the host social structure available to rabies virus. We know that the social systems of vampire bats can be used to advantage by humans for spreading fatal anticoagulants (e.g., diphenadiane) or vaccines in the wild (Bakker et al. 2019).
Bats may be labelled by their choice of roosts. In Canada, house bats may roost in buildings (e.g., Myotis spp. and big brown bats), whereas tree-roosting bats do not (e.g., eastern red and hoary bats). As previously mentioned, silver-haired bats rarely enter or roost in buildings. Arguably, humans are more likely to encounter bats that roost in buildings. This leaves us with a conundrum: three of the last four cases of human rabies acquired in Canada from a bat involved the silver-haired bat rabies virus variant (Table 1). The transmission of this variant among bat species may help to explain this situation (Nadin-Davis et al. 2001). In other words, a little brown myotis could be infected by the silver-haired bat rabies virus variant.
Recommendations
The combination of realities we have outlined makes it imperative for people to avoid being bitten by bats. Biologists and others that regularly contact and handle bats should receive a rabies pre-exposure immunization series and have their protective antibody titre measured regularly, every 6 to 24 months (depending on the profession or activities), and obtain booster doses of vaccine as needed. Wherever possible, only biologists or other trained wildlife professionals should handle bats. Humans bitten by a bat should immediately wash and thoroughly flush and clean the wound with soap and water, and then immediately report the incident to a local Medical Officer of Health or a Canadian Food Inspection Agency (CFIA) veterinarian who can help to arrange appropriate PEP. This recommendation also applies to bites from other wild or feral mammals. In the case of bat bites, the bat should be contained so a trained professional can euthanize it and submit the body for rabies testing and species identification.
Two other situations are grim reminders of possible hazards involving humans and bat variants of the rabies virus. First, transmission to humans has occurred through accidents in the laboratory associated with the aerosolization of concentrated virus (Winkler et al. 1973; Tillotson et al. 1977). Second, and more problematic, are reports of people being exposed to rabies virus through the air in caves harbouring millions of bats (Constantine 1962). Either situation makes it impossible to categorically deny the possibility of aerosol transmission even though it remains doubtful that this occurs commonly (i.e., it remains a possibility in very rare circumstances). Perhaps fortunately, the very high densities of bats observed in some places, like the ∼20 million Brazilian free-tailed bats that roost in Bracken Cave, Texas, USA, do not occur in Canada.
Some reflections
Associating bats with frightening zoonotic diseases adversely affects both the public image and conservation of bats (Tuttle 2017). When it comes to public health, a negative image associated with the ∼1400 species of bats reflects a mixture of truth and misinformation. Bats are long-term reservoirs for rabies virus and we have explored this situation. Some other assertions about bats and diseases are not based on strong evidence. For example, Yang et al. (2019) identified an Ebola-like virus associated with bats. The virus was isolated from a liver sample from one fruit bat in China, and the bat was not identified beyond the level of genus (Rousettus). What does this prove about bats or Ebola? Sample sizes of n = 1 are worrisome. Some researchers may have been utterly mislead in tying bats to Middle East respiratory syndrome (MERS) based on data from one bat (Memish et al. 2013). Careful research about viruses in bats is one thing, but assertions that bats are long-term reservoir hosts for feared diseases such as Ebola (e.g., Calisher et al. 2006) or severe acute respiratory syndrome (SARS) (Racey et al. 2018) are another. Too often assertions are perpetuated as truths with little regard for actual evidence and (or) context (e.g., “bat flu”—Karakus et al. 2019).
As noted above, we typically lack important details about situations where humans are exposed to rabid bats. Take Brazil, for example. There Schneider et al. (1996) interviewed 129 of 160 people from one remote Amazonian village to assess the incidence of vampire bats feeding on humans. In this and other villages, vampire bats feed on humans and the incidence of bites can vary widely from 23% through 41%–88% of residents (Fenton et al. 2020). It is difficult to put these numbers in a broader perspective because according to the World Health Organization there were three human deaths from rabies in Brazil in 2016 and all involved bats (paho.org/salud-en-las-americas-2017/?tag=rabies). For Latin America, Escobar et al. (2015) reported 0.00–0.10 human deaths from rabies per 100 000 in the population. In the final analysis, the chances of any one person contracting rabies from a vampire bat are exceedingly low (Fenton et al. 2020). In short, people should neither dismiss the possibility of rabies associated with a bite by a wild bat nor use fear of rabies to label all bats as a looming threat to humans.
Acknowledgements
Our research on bats and (or) rabies has been generously supported by the Natural Sciences and Engineering Council of Canada and the Canadian Institutes of Health Research. We thank current and former students and post-doctoral fellows for their observations over the years, and K. Delaney and the staff of the Psychology Animal Facility at McMaster University for animal care and support. We thank two anonymous reviewers for their comments about an earlier version of this manuscript.
References
Baer GM, and Adams DB. 1970. Rabies in insectivorous bats in the United States, 1953–65. Public Health Reports, 85: 637–645.
Baer GM, and Bales GL. 1967. Experimental rabies infection in the Mexican freetail bat. Journal of Infectious Diseases, 117: 82–90.
Bakker KM, Rocke TE, Osorio JE, Abbott RC, Tello C, Carrera JE, et al. 2019. Fluorescent biomarkers demonstrate prospects for spreadable vaccines to control disease transmission in wild bats. Nature Ecology & Evolution, 3: 1697–1704.
Bell GP. 1980. A possible case of interspecific transmission of rabies in insectivorous bats. Journal of Mammalogy, 61: 528–530.
Bradley JA. 1979. Laboratory diagnosis of rabies in western Canada (1968–1977). The Canadian Veterinary Journal, 20: 186–190.
Brass DA. 1994. Rabies in bats: natural history and public health consequences. Livia Press, New Haven, Connecticut.
Brown S. 2019. B.C. man dies of rabies after contact with a bat on Vancouver Island. Vancouver Sun, 16 July [online]: Available from vancouversun.com/news/local-news/b-c-man-dies-of-rabies-after-contact-with-a-bat-on-vancouver-island.
Calisher CH, Childs JE, Field HE, Holmes KV, and Schountz T. 2006. Bats: important reservoir hosts of emerging viruses. Clinical Microbiology Reviews, 19: 531–545.
Constantine DG. 1962. Rabies transmission by nonbite route. Public Health Reports, 77: 287–289.
Constantine DG. 1967. Bats in the southwestern United States. Public Health Reports, 82: 867–888.
Constantine DG. 1971. Bats in relation to the health, welfare and economy of man. In Biology of bats. Edited by WA Wimsatt. Academic Press, New York, New York. Vol. 2, pp. 319–449.
Constantine DG. 2009. Bat rabies and other lyssavirus infections. U.S. Geological Survey Circular 1329. U.S. Geological Survey, Reston, Virginia. 68 p.
Constantine DG, Tierkel ES, Kleckner MD, and Hawkins DM. 1968. Rabies in New Mexico cavern bats. Public Health Reports, 83: 303–316.
Courter RD. 1954. Bat rabies. Public Health Reports, 69: 9–16.
De Serres G, Dallaire F, Côte M, and Skowronski DM. 2008. Bat rabies in the United States and Canada from 1950 through 2007: human cases with and without bat contact. Clinical Infectious Diseases, 46: 1329–1337.
De Serres G, Skowronski DM, Mimault P, Ouakki M, Maranda-Aubut R, and Duval B. 2009. Bats in the bedroom, bats in the belfry: reanalysis of the rationale for rabies post-exposure prophylaxis. Clinical Infectious Diseases, 48: 1493–1499.
Dempster G, Grodums EI, Bayatpour M, and Zbitnew A. 1972. A human case of unsuspected rabies in Saskatchewan diagnosed by virus isolation. Canadian Journal of Public Health, 63: 215–218.
Dermaux-Msimang V, Blumberg L, Paweska J, and Weyer J. 2019. Epidemiology of human rabies in South Africa. NCID Bulletin, 17: 63–72.
Despond O, Tucci M, Decaluwe H, Gregoire M-C, Teitelbaum JS, and Turgeon N. 2002. Rabies in a nine-year-old child: the myth of the bite. Canadian Journal of Infectious Diseases and Medical Microbiology, 13: 121–125.
Dolman CL, and Charlton KM. 1987. Massive necrosis of the brain in rabies. Canadian Journal of Neurological Sciences, 14: 162–165.
Dyer JL, Wallace R, Orciari L, Hightower D, Yager P, and Blanton JD 2013. Rabies surveillance in the United States during 2012. Journal of the American Veterinary Medical Association, 243(6): 805–815.
Editorial. 2019. Build trust to combat Ebola. Nature, 567: 433.
Elmgren LD, Nadin-Davis SA, Muldoon FT, and Wandeler AI. 2002. Diagnosis and analysis of a recent case of human rabies in Canada. Canadian Journal of Infectious Diseases, 13: 129–133.
Escobar LE, Peterson AT, Favi M, Yung V, and Merdina-Vogel G. 2015. Bat-borne rabies in Latin America. Revista do Instituto de Medicina Tropical de São Paulo, 57: 63–72.
Fenton MB, and Simmons NB. 2015. Bats: a world of science and mystery. University of Chicago Press, Chicago, Illinois.
Fenton MB, Streicker DG, Racey PA, Tuttle MD, Medellin RA, Daley MJ, et al. 2020. Knowledge gaps about rabies transmission from vampire bats to humans. Nature Ecology & Evolution, 4: 517–518.
Fenton MB, Jackson AC, and Faure PA. In press. Bat rabies in Canada. In Taking the bite out of rabies: the evolution of rabies management in Canada. Edited by DJ Gregory and RR Tinline. University of Toronto Press, Toronto, Ontario.
Fooks AR, and Jackson AC (Editors). 2020. Rabies: scientific basis of the disease and its management. 4th edition. Elsevier Academic Press, London, UK.
Franka R, Johnson N, Müller T, Vos A, Neubert AL, Freuling C, et al. 2008. Susceptibility of North American big brown bats (Eptesicus fuscus) to infection with European bat lyssavirus type 1. Journal of General Virology, 89: 1998–2010.
George DB, Webb CT, Farnsworth ML, O’Shea TJ, Bowen RA, Smith DL, et al. 2011. Host and viral ecology determine bat rabies seasonality and maintenance. Proceedings of the National Academy of Sciences of the United States of America, 108: 10208–10213.
Hampson K, Coudeville L, Lembo T, Sambo M, Kieffer A, Attlan M, et al. 2015. Estimating the global burden of endemic canine rabies. PLoS Neglected Tropical Diseases, 9: e0003709.
Huot C, De Serres G, Duval B, Maranda-Aubut B, Ouakki M, and Skowronski DM. 2008. The cost of preventing rabies at any cost: post-exposure prophylaxis for occult bat contact. Vaccine, 26: 4446–4450.
Jackson AC. 2011a. Update on rabies. Research and Reports in Tropical Medicine, 2: 31–43.
Jackson AC. 2011b. Rabies in the critical care unit: diagnostic and therapeutic approaches. Canadian Journal of Neurological Sciences, 38: 689–695.
Jackson AC. 2020a. Human disease. In Rabies: scientific basis of the disease and its management. Edited by AR Fooks and AC Jackson. 4th edition. Elsevier Academic Press, London, UK.
Jackson AC. 2020b. Pathogenesis. In Rabies: scientific basis of the disease and its management. Edited by AR Fooks and AC Jackson. 4th edition. Elsevier Academic Press, London, UK.
Jackson AC, and Fenton MB. 2001. Human rabies and bat bites. The Lancet, 357: 1714.
Johnstone J, Saxinger L, McDermid R, Bagshaw S, Resch L, Lee B, et al. 2008. Human rabies—Alberta, Canada, 2007. Morbidity and Mortality Weekly Report, 57: 197–200.
Karakus U, Thamamongood T, Ciminski K, Ran W, Gunther SC, Pohl MO, et al. 2019. MHC Class II proteins mediate cross-species entry of bat influenza viruses. Nature, 567: 109–112.
King DB, Sangalang VE, Manuel R, Marrie T, Pointer AE, and Thomson AD. 1978. Suspected case of human rabies—Nova Scotia. Canadian Disease Weekly Report, 4: 49–51.
Klug BJ, Turmelle AS, Ellison JA, Baerwald EF, and Barclay RMR. 2011. Rabies prevalence in migratory tree-bats in Alberta and the influence of roosting ecology and sampling method on reported prevalence of rabies in bats. Journal of Wildlife Diseases, 47: 64–77.
Kuzmin IV, and Rupprecht CE. 2007. Bat rabies. In Rabies. Edited by AC Jackson and WH Wunner. Elsevier Academic Press, London, UK. pp. 259–307.
Mani RS, Anand AM, and Madhusudana SN. 2016. Human rabies in India: an audit from a rabies diagnostic laboratory. Tropical Medicine and International Health, 21: 556–563.
McDermid RC, Saxinger L, Lee B, Johnstone J, Noel Gibney RT, Johnson M, et al. 2008. Human rabies encephalitis following bat exposure: failure of therapeutic coma. Canadian Medical Association Journal, 178: 557–561.
Memish ZA, Mishra N, Olival KJ, Fagbo SF, Kapoor V, Epstein JH, et al. 2013. Middle East respiratory syndrome coronavirus in bats, Saudi Arabia. Emerging Infectious Diseases, 19(11): 1819–1823.
Moore GJ, and Raymond RJ. 1970. Prolonged incubation period of rabies in a naturally infected insectivorous bat, Eptesicus fuscus (Beauvois). Journal of Wildlife Diseases, 6: 167–168.
Moreno JA, and Baer GM. 1980. Experimental rabies, in the vampire bat. American Journal of Tropical Medicine and Hygiene, 29: 254–259.
Nadin-Davis SA, Huang W, Armstrong J, Casey GA, Bahloul C, Tordo N, et al. 2001. Antigenic and genetic divergence of rabies viruses from bat species indigenous to Canada. Virus Research, 74: 139–156.
Nadin-Davis SA, Feng Y, Mousse D, Wandeler AI, and Aris-Brosou S. 2010. Spatial and temporal dynamics of rabies virus variants in big brown bat populations across Canada: footprints of an emerging zoonosis. Molecular Ecology, 19: 2120–2136.
National Advisory Committee on Immunization. 1998. Canada immunization guide. 5th edition. Canadian Medical Association, Ottawa, Ontario.
National Advisory Committee on Immunization. 2002. Canada immunization guide. 6th edition. Canadian Medical Association, Ottawa, Ontario.
National Advisory Committee on Immunization. 2009. Recommendations regarding the management of bat exposures to prevent human rabies. Canada Communicable Disease Report, 35(ACS-7): 1–28.
Parker RD, McKay D, Hawes C, Daly P, Bryce E, Doyle P, et al. 2003. Human rabies, British Columbia—January 2003. Canada Communicable Disease Report, 29: 137–138.
Picard AC. 1984. Human rabies acquired outside of Canada—Quebec. Canadian Diseases Weekly Report, 10: 177–178.
Prins B, and Loewen KG. 1991. Rabies diagnosis in western Canada, 1985–1989. The Canadian Veterinary Journal, 32: 487–491.
Racey PA, Fenton MB, and Mubareka S. 2018. Don’t misrepresent link between bats and SARS. Nature, 553: 281.
Ripperger S, Gunther L, Wieser H, Duda N, Hierold M, Casens B, et al. 2019. Proximity sensors on common noctule bats reveal evidence that mothers guide juveniles to roosts but not food. Biology Letters, 15: 20180884.
Rosatte RC. 1987. Bat rabies in Canada: history, epidemiology and prevention. The Canadian Veterinary Journal, 28: 754–756.
Rossiter JP, and Jackson AC. 2020. Pathology. In Rabies: scientific basis of the disease and its management. Edited by AR Fooks and AC Jackson. 4th edition. Elsevier Academic Press, Oxford, UK.
Rupprecht CE, Hanlon CA, and Hemachudha T. 2002. Rabies re-examined. The Lancet Infectious Diseases, 2: 327–343.
Sadler WW, and Enright JB. 1959. Effect of metabolic level of the host upon the pathogenesis of rabies in the bat. Journal of Infectious Diseases, 105: 267–273.
Schneider MC, Santos-Burgoa C, Aron J, Mnoz B, Ruiz-Velazco S, and Uieda W. 1996. Potential force of infection of human rabies transmitted by vampire bats in the Amazonian Region of Brazil. American Journal of Tropical Medicine and Hygiene, 55: 680–684.
Schowalter DB. 1980. Characteristics of bat rabies in Alberta. Canadian Journal of Comparative Medicine, 44: 70–76.
Shankar V, Bowen RA, Davis AD, Rupprecht CE, and O’Shea TJ. 2004. Rabies in a captive colony of big brown bats (Eptesicus fuscus). Journal of Wildlife Diseases, 40: 403–413.
Sulkin SE, Allen R, Sims R, Krutzsch PH, and Kim C. 1960. Studies on the pathogenesis of rabies in bats. II. Influence of environmental temperature. Journal of Experimental Medicine, 112: 595–617.
Tillotson JR, Axelrod D, and Lyman DO. 1977. Rabies in a laboratory worker—New York. Morbidity and Mortality Weekly Report, 26: 183–184.
Turgeon N, Tucci M, Teitelbaum J, Deshaies D, Pilon PA, Carsley J, et al. 2001. Human rabies—Quebec, Canada, 2000. Journal of the American Medical Association, 285: 160–161.
Tuttle MD. 2017. Give bats a break. Issues in Science and Technology, 33: 41–50.
Van Zyll de Jong CG. 1985. Handbook of Canadian mammals: volume 2. Bats. National Museum of Natural Science, National Museums of Canada, Ottawa, Ontario.
Vargas AM, Romano APO, and Merchán-Hamann E. 2019. Human rabies in Brazil: a descriptive study, 2000–2017. Epidemiologia e Serviços de Saúde, 28: e2018275.
Walker G, Thiessen B, Graeb D, Wayne Moore GR, and Mackenzie IRA 2016. An unusual case of rabies encephalitis. Canadian Journal of Neurological Sciences, 43(6): 852–855.
Warshawsky B, and Desai S. 2010. Exposure to bats: updated recommendations. Canadian Medical Association Journal, 182: 60.
Webster WA, Casey GA, Charlton KM, Picard AC, and McLaughlin B. 1985. Human rabies acquired outside of Canada. Canada Disease Weekly Report, 11: 13–14.
Webster WA, Casey GA, Charlton KM, Sayson RC, McLaughlin B, and Noble MA. 1987. A case of human rabies in western Canada. Canada Journal of Public Health, 78: 412–413.
Wilcox ME, Poutanen SM, Krajden S, Agid R, Kiehl TR, and Tang-Wai DF. 2014. A case of rabies encephalitis: new dog, old tricks. Neurology, 82(12): 1101.
Willis CKR, and Brigham RM. 2007. Social thermoregulation exerts more influence than microclimate on forest roost preferences by a cavity-dwelling bat. Behavioral Ecology and Sociobiology, 62: 97–108.
Winkler WG, Fashinell TR, Leffingwell L, Howard P, and Conomy JP. 1973. Airborne rabies transmission in a laboratory worker. Journal of the American Medical Association, 226: 1219–1221.
Yang X-L, Tan CW, Anderson DE, Jiang R-D, Li B, Zhang W, et al. 2019. Characterization of a filovirus (Měnglà virus) from Rousettus bats in China. Nature Microbiology, 4: 390–395.
Information & Authors
Information
Published In

FACETS
Volume 5 • Number 1 • January 2020
Pages: 367 - 380
Editor: Mark Mallory
History
Received: 18 December 2019
Accepted: 20 March 2020
Version of record online: 1 June 2020
Copyright
© 2020 Fenton et al. This work is licensed under a Creative Commons Attribution 4.0 International License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Data Availability Statement
All relevant data are within the paper.
Key Words
Sections
Subjects
Authors
Author Contributions
All conceived and designed the study.
All analyzed and interpreted the data.
All contributed resources.
All drafted or revised the manuscript.
Competing Interests
The authors have declared that no competing interests exist.
Metrics & Citations
Metrics
Other Metrics
Citations
Cite As
M. Brock Fenton, Alan C. Jackson, and Paul A. Faure. 2020. Bat bites and rabies: the Canadian scene. FACETS.
5(1): 367-380. https://doi.org/10.1139/facets-2019-0066
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
Cited by
1. Bat handlers, bat bites, and rabies: vaccination and serological testing of humans at risk
2. A cross-sectional survey on fruit bat-human interaction in Pakistan; one health perspective
3. The Ecological Range and Principles of Wildlife Rabies Virus Perpetuation in the Americas
4. Assessing the extent and public health impact of bat predation by domestic animals using data from a rabies passive surveillance program
5. Urban Bats, Public Health, and Human-Wildlife Conflict