Spatiotemporal patterns of low and highly pathogenic avian influenza virus prevalence in murres in Canada from 2007 to 2022—a case study for wildlife viral monitoring
Abstract
Migratory seabirds move across ocean basins and are one of the primary reservoirs of low pathogenic avian influenza virus (LPAIV). This includes the millions of thick-billed murres (Uria lomvia) and common murres (Uria aalge) that are distributed across northern hemisphere oceans. In response to increasingly frequent detections of highly pathogenic avian influenza virus (HPAIV) in Europe in 2020–2021, avian influenza virus (AIV) monitoring in wildlife has increased. We compiled data from murres tested for AIV in Canada between 2007 and 2022 to quantify spatiotemporal variation in the prevalence of LPAIV and HPAIV in these birds. No HPAIV was detected in murres prior to 2022, but HPAIV was present in 46% of both live/harvested and found dead murres in the northwestern Atlantic in 2022 with prevalence peaking at 63% among live birds in the summer. In the eastern Canadian Arctic, HPAIV prevalence in 2022 was <1% while LPAIV prevalence was 21%, which was significantly higher than previous sampling years. Power analyses suggest approximately 100 samples from breeding murres should be collected annually per colony or region to detect moderate changes in HPAIV prevalence. These analyses inform robust monitoring of viruses in wildlife, with implications for conservation, harvest management, and public health.
Introduction
Wild birds are natural reservoirs of avian influenza virus (AIV), but until 2005 they appeared to be mostly refractory to serious disease caused by infection with highly pathogenic AIV (HPAIV; Liu et al. 2005). However, since 2022, frequent outbreaks, high mortality, and intercontinental spread of HPAIV subtype H5N1 lineage A/goose/Guangdong/1/1996 (Gs/GD) clade 2.3.4.4b have made wild birds a focal point for HPAIV wildlife monitoring worldwide (Ramey et al. 2022; Harvey et al. 2023).
Increased detection of HPAIV-attributable morbidity and mortality in poultry, wild birds, free-living mesocarnivores, and marine mammals in recent years has sparked global concern (Verhagen et al. 2021; Alkie et al. 2022, 2023; Banyard et al. 2022; Caliendo et al. 2022). In Eurasia, HPAIVs are considered to be enzootic, circulating endemically in wild birds (Verhagen et al. 2021; Ramey et al. 2022). The recent changes in HPAIV dynamics in wildlife are due to the emergence of HPAIV H5 clade 2.3.4.4b, which appeared in 2014, and now dominates HPAI disease burden globally with sustained transmission in wild birds (Verhagen et al. 2021). Mass mortality events attributed to 2.3.4.4b have been reported in great skuas (Stercorarius skua) in Scotland (Banyard et al. 2022), South American sea lions (Otaria flavescens) in Peru (Gamarra-Toledo et al. 2023), and harbour seals (Phoca vitulina) in North America, among many species (NOAA 2022). Notably, in 2022, seabirds (e.g., auks, sulids, larids) were particularly affected by 2.3.4.4b, which led to unusual mass mortality events in both North America and Europe (USGS 2022; WHISPers, 2023). Despite global efforts to track and understand the H5 2.3.4.4b clade, it is unclear how common HPAIV infection is among migratory seabirds (Lang et al. 2016).
Murres are a genus of seabirds—also known as guillemots in Europe, turrs in Newfoundland and Labrador, Canada, and as akpa (⊲b∠) in Nunavut, Canada—that includes common murre (COMU; Uria aalge) and thick-billed murre (TBMU; Uria lomvia). They are abundant, migrate across ocean basins (Frederiksen et al. 2016), and congregate at high-density breeding sites, with potential for high levels of fecal-oral and respiratory transmission of viruses at their colonies (Huang et al. 2014). Murres have been identified as reservoir hosts of low pathogenic AIV (LPAIV) throughout their range in the northern hemisphere (Granter et al. 2010; Huang et al. 2014; Wille et al. 2014; Lang et al. 2016), and have now also been found to be infected by HPAIV in the Netherlands (Caliendo et al. 2022) and Scotland (Falchieri et al. 2022). In Canada, AIV data from murres are available from 2007–2022, before and since 2021 HPAIV incursion(s) in wild birds of North America.
Most wildlife monitoring programs in Canada in 2022 were risk-based, with planning based on known information on AIV in some species. However, this study explores the use of AIV monitoring data at a species-level and how its findings can inform hunters on harvest limits and provide them with guidance regarding risk mitigation and education campaigns. Ideally, disease monitoring is a component of broader population models of mortality and morbidity from multiple sources, which feed into conservation and management priorities and guided actions (Fig. 1). Ongoing monitoring in wild birds and commercial poultry will help identify deviations from normal disease circulation and burden.
Fig. 1.
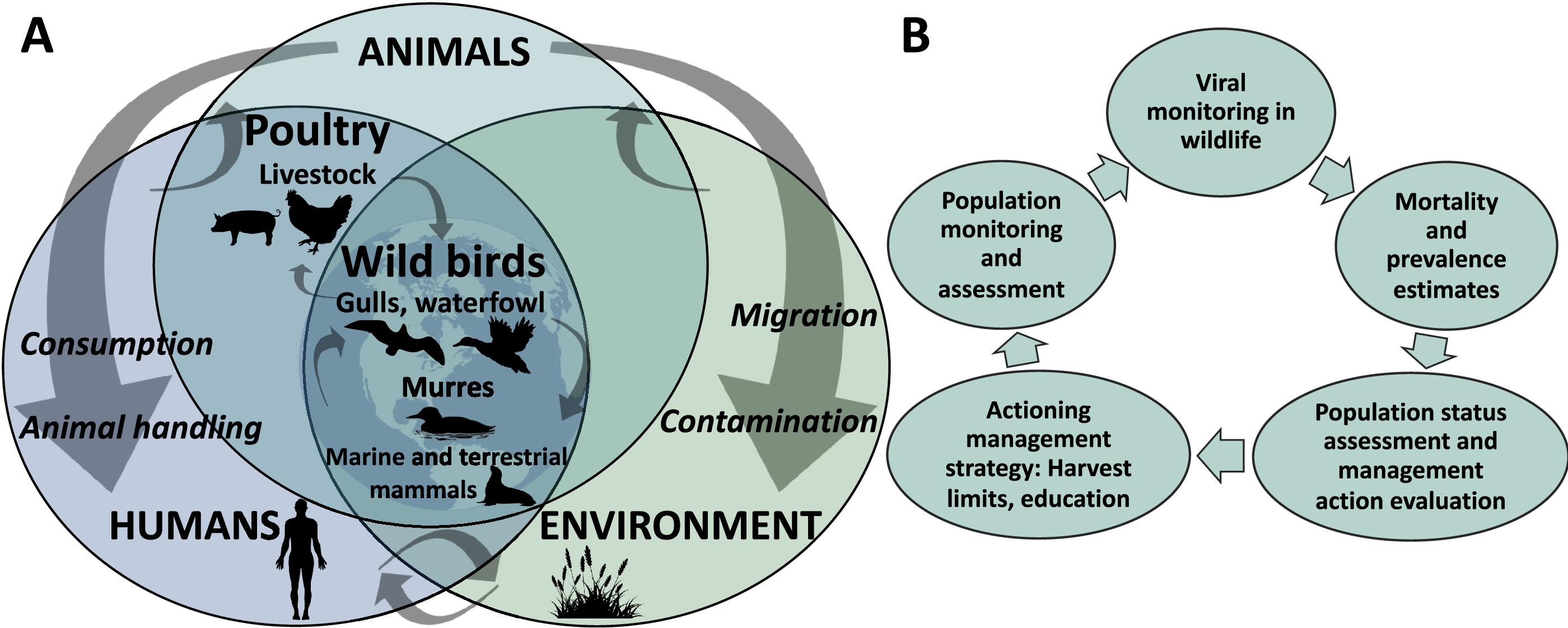
Given this context, this analysis is a case study of monitoring viral prevalence in wildlife within a regional outbreak with elevated urgency in a species with conservation, management, and public health implications (Stephen et al. 2018). Prevalence of HPAIV and LPAIV infection were calculated in murres sampled across Canada in 2022, and compared to 2007–2021, where monitoring was episodic as part of long-term population monitoring and research priorities. Our objectives were to: (i) report the detection of HPAIV in murres in Canada in 2022 since the HPAIV incursions into North America; (ii) evaluate differences in LPAIV and HPAIV prevalence in murres in 2022 across regions and sampling months; (iii) investigate how AIV prevalence has changed in murres in Canada in 2022 compared to previous years; and (iv) apply power analyses to inform sampling plans for future viral monitoring in relation to the wildlife health policy needs for murres. Further, we discuss the successes and areas for improvement in the monitoring of pathogens in wildlife generally.
Methods
Data sources
This study used data collected across multiple Canadian provinces and territories from the Atlantic, Arctic, and Pacific marine ecosystems. Oral and cloacal swab samples were collected from live/harvested murres in Nunavut (NU) in the eastern Arctic and Newfoundland and Labrador (NL) in the Northwest Atlantic, and from murres found dead due to any cause or found sick in NL, New Brunswick (NB), Nova Scotia (NS), Prince Edward Island (PEI), Quebec (QC) (all Northwest Atlantic), and British Columbia (BC) (eastern North Pacific).
The dataset was compiled from the Canadian Wildlife Health Cooperative (CWHC), Environment and Climate Change Canada (ECCC), and provinces and territories (P/Ts), and included data previously published for samples from NL between 2008 and 2011 (Granter et al. 2010; Huang et al. 2014; Wille et al. 2014), yet-unpublished data from NU from 2011 to present (Provencher et al. in prep.), unpublished historic data, and contemporary 2022 data. Data sources were summarized for 2007–2021 by the number of samples by initial testing laboratories (Table S1), confirmatory diagnostic laboratories (Table S2), and submitting agencies (Table S3), and for the 2022 data, by preliminary testing labs (Table S4) and ECCC project names (Table S5).
Sample collection and laboratory analysis
Although capture and sample collection protocols varied somewhat across sites and personnel, swabs were collected from murres captured live and released, harvested, or found dead or sick. Oral and cloacal swabs from a given individual were often pooled in the same universal transport medium vial. CWHC manages a passive surveillance program for AIV, whereby submitted carcasses are routinely necropsied, swabbed, and tested for AIV. In 2022, due to the unusually high number of mass mortality events, a field-level triage protocol was put in place to limit redundant testing of dead birds, including murres. Therefore, samples represent a subsample of a larger group of found dead murres. Live murres at breeding colonies in NU and NL were often swabbed in conjunction with ongoing research efforts focused on murre populations (e.g., tracking studies, banding programs), as described for the 2022 data in Tables S4–S5. Murres are annually harvested in NU and NL by Indigenous and non-Indigenous hunters, and thus samples from harvested birds were obtained through partnerships with Indigenous governments and organizations in these regions (e.g., Nunatsiavut Government, Nattivak Hunter and Trapper Organization, Sululiit Area Co-Management Committee, and Mittimatalik Hunters and Trappers Organization). Samples from non-Indigenous hunters in NL were obtained opportunistically from individual hunters willing to have their murres sampled, and from murres seized and forfeited to ECCC enforcement officers.
Samples from murres collected in Nunavut in 2007–2021 were primarily sent to the Animal Health Laboratory (AHL) at the University of Guelph, while samples from 2022 were processed by AHL, Memorial University of Newfoundland (MUN), and the National Centre for Foreign Animal Disease (NCFAD). Handling of bird samples at MUN was under protocol 20-05-AL from the Institutional Animal Care Committee and Biosafety Permit S-103. At AHL and MUN, initial screening was conducted by real-time reverse transcriptase polymerase chain reaction (RT-PCR) targeting the AIV matrix (M) gene. Any non-negative samples were further tested by RT-PCR for presence of H5 and H7 subtype viruses. All H5 non-negative samples were sent to the NCFAD for confirmatory testing, isolation, and genomic sequencing. Non-negative includes positive (cycle threshold, Ct < 36) and inconclusive (36 ≤ Ct ≤ 40) results (CWHC 2022). Thresholds for positive and inconclusive results were in alignment with the standard applied by the NCFAD reference laboratory and used by all laboratories in the Canadian Animal Health Surveillance Network (CAHSN), based on prior literature (Spackman et al. 2002; Granter et al. 2010; Wille et al. 2014). A previous study from which data are included here assigned samples with Ct < 35 as positives (Wille et al. 2014).
Data curation
Contemporary data for 2022 were downloaded from the CWHC database on 22 November 2022 and joined with additional datasets of hunter-harvested murres from NL provided on 23 December 2022, and murres in BC provided on 27 August 2023. Samples were inspected for duplicate IDs and complete sample dates. Metadata included species (COMU, TBMU), status (live/harvested; found sick or found dead), latitude/longitude (lat/long), province, age class (after hatch year or adult, AHY; hatch year or juvenile, HY), sex (only discernible for murres found dead), capture method, RT-PCR results for M, H5, and H7 genes, lineage assignment to 2.3.4.4b, and origin of genome segments as Eurasian or North American based on whole-genome sequencing (Giacinti et al. 2023). Samples where RT-PCR M results were missing were removed from the analysis (n = 1).
Data from 2007 to 2021 were downloaded from the CWHC database on 25 November 2022 and combined with the BC data. Geographic metadata (province, region) were incomplete, therefore lat/long values were used to identify the region of sampling and to compare differences among sampling sites. Two samples had unrealistic lat/long values of “0, −1”. In the first instance, the sampling city was provided and that was used to ascribe a lat/long based on other samples from that city. In the second instance, only province was provided and it was ascribed the lat/long most commonly sampled in the province within two days of the sample. Five samples from Prince Rupert, BC had no sample collection date, only year; since their causes of death were gillnet entrapment, their dates were estimated to have occurred during the Nass and Skeena sockeye salmon (Oncorhynchus nerka) runs in mid-July 2010. Duplicate band IDs were excluded if they were collected in the same sampling month and year (n = 12, including: in Nunavut, n = 2 from 2009 and n = 8 from 2011; in Newfoundland, n = 1 from 2009 and n = 1 from 2011). Samples with missing RT-PCR results for M were removed (n = 171).
Statistical analyses
To compare regional patterns, samples from COMU and TBMU were grouped together; however, the analyses were stratified by bird status: live/harvested or found dead (or sick). Data from NL, NB, NS, PEI, and QC were grouped together as the Atlantic region, while data from BC is referred to as the Pacific region, and NU is referred to as the Arctic region.
Viral period (average) prevalence was calculated by status, year, season (breeding summer months, June–August, vs. rest of year), month, region, province/territory, and site. For murres found dead, the terminology “percentage positivity” is utilized instead of “prevalence” because cause of death was not always known and birds found dead were not representative of the wider population. The nomenclature AIV, LPAIV, and HPAIV are used throughout to denote the presence of virus, whereas we did not investigate the presence of disease, denoted by LPAI and HPAI. AIV positivity (% AIV+) was estimated as the number of samples with non-negative M tests divided by the total number of samples tested, per hundred (Fig. S1). HPAIV positivity (% HPAIV+) was estimated as the number of samples with non-negative M and H5 tests divided by the total number of samples tested, per hundred. LPAIV positivity (% LPAIV+) was estimated as the number of samples with non-negative M but negative H5 tests divided by the total number of samples tested, per hundred. Proportions with 95% confidence intervals were calculated using one group two-sided proportion tests.
Statistical comparisons of the AIV, HPAIV, and LPAIV prevalence across time and space were conducted using two-sided χ2 tests with Yates continuity correction. Fisher’s exact tests were used if any of the assumptions for a χ2 test were violated, including that >80% of expected frequencies exceed 5 and all expected frequencies exceed 1 (McHugh 2013). Statistical tests were summarized by p-value and power to detect a true association of the estimated effect size (Table S6). Binomial generalized linear models with a logit link function were built separately by status to test the association of the response variables, AIV or HPAIV positivity, with the primary predictor, sampling region, considering inclusion of potential confounding variables, season, species, and age category. Likelihood ratio tests were used to test whether confounding variables improved the goodness-of-fit using forward stepwise selection. Final models were interpreted as adjusted odds ratios.
Sample sizes to target for future monitoring years were calculated using two-sided two-proportions with unequal sample size power analyses assuming the baseline comparison to the % AIV+ or % HPAIV+ among live birds (mean and confidence limits) and number sampled in 2022, with a range of expected changes (primary analysis: 25% increase), resultant effect sizes, power values (50%, 80%), and alpha significance values (0.01, 0.05). R packages used in the analysis are listed in the supplementary materials.
Mapping
Cartographic boundary files were obtained for Canadian provinces from the 2016 Census as NAD83 Lambert datum (EPSG:3348) (Statistics Canada 2019), for the United States as WGS84 (United States Census Bureau 2019), and for Greenland as WGS84 (gadm.org 2022). The US and Greenland were reprojected as EPSG:3348 in units of meters. All lat/long values in decimal degrees were truncated to three decimal points for site-specific confidentiality, assigned the WGS84 (EPSG:4326) coordinate reference system, then reprojected to EPSG:3348. R packages used to generate maps are listed in the supplementary materials.
Results
HPAIV and LPAIV detected among Canadian murres in 2022
There were 427 murres tested for AIV in 2022 across Canada, including 305 live/harvested birds and 122 birds found dead (Table 1). Live/harvested birds consisted mostly of TBMU (n = 260, 85.2%), which were sampled in the Arctic via research programs (n = 248, 81.3%) and in NL within the Atlantic region via hunter-based collections (n = 57, 18.7%). Dead birds were mostly COMU (n = 81, 66.4%) and were most widely sampled in the Atlantic where they would have been local breeders (n = 109, 89.3%), with additional sampling in the Pacific (n = 13, 10.7%). Among birds where age group was recorded, 96.8% (n = 212) of live murres were after-hatch year (AHY) and 73.3% (n = 22) of dead murres were AHY (though age data were mostly missing). Sex in murres can only be determined by internal examination or deoxyribonucleic acid, so was not available for live birds; 61.1% (n = 11) of dead birds where sex was determined were male. Live birds were primarily sampled in July (n = 165, 54.1%) and August 2022 (n = 124, 40.7%), while samples from dead birds were dispersed through the year (Fig. 4).
Table 1.
| Category | Group | Live/harvested, N (%) | Dead, N (%) |
|---|---|---|---|
| Total | 305 (100) | 122 (100) | |
| Species | Common murres (COMU) | 45 (14.8) | 81 (66.4) |
| Thick-billed murres (TBMU) | 260 (85.2) | 41 (33.6) | |
| Age group | After hatch-year (AHY) | 212 (96.8) | 22 (73.3) |
| Hatch-year (HY) | 7 (3.2) | 8 (26.7) | |
| Unknown | 9 | 92 | |
| Sex | Female | – | 7 (38.9) |
| Male | – | 11 (61.1) | |
| Unknown | 305 | 104 | |
| Region | Pacific | – | 13 (10.7) |
| Arctic | 248 (81.3) | – | |
| Atlantic | 57 (18.7) | 109 (89.3) | |
| Atlantic: Newfoundland and Labrador (NL) | 57 (18.7) | 82 (67.2) | |
| Atlantic: Nova Scotia (NS) | – | 16 (13.1) | |
| Atlantic: New Brunswick (NB) | – | 4 (3.3) | |
| Atlantic: Prince Edward Island (PEI) | – | 4 (3.3) | |
| Atlantic: Quebec (QC) | – | 3 (2.5) | |
| Month | Jan 2022 | 3 (1.0) | 1 (0.8) |
| Feb 2022 | – | 6 (4.9) | |
| Mar 2022 | – | 20 (16.4) | |
| Apr 2022 | – | 28 (23.0) | |
| May 2022 | – | 1 (0.8) | |
| Jun 2022 | – | 11 (9.2) | |
| Jul 2022 | 165 (54.1) | 28 (23.0) | |
| Aug 2022 | 124 (40.7) | 25 (20.5) | |
| Sep 2022 | – | 1 (0.8) | |
| Oct 2022 | – | – | |
| Nov 2022 | – | 1 (0.8) | |
| Dec 2022 | 13 (4.2) | – |
Note: Percentages by category were calculated excluding missing data.
RT-PCR results were summarized as AIV− (M−), LPAIV+ (M+, H5−, or H5 test unavailable), or HPAIV+ (M+, H5+) (Fig. S1; Fig. 3). There were no detections of H7 in the murres. Our presumption that all the H5+ samples were HPAIV was corroborated by the fact that among the 35 H5+ samples that had their whole genomes sequenced (all from dead COMU), all were within H5N1 clade 2.3.4.4b, of which 33 were inferred to have fully Eurasian lineage genomes, while 2 samples were reassortants with 50% of segments (HA, NA, M, and NS) from Eurasian lineages and 50% of segments (NP, PB1, PB2, and PA) from North American lineages (Fig. S3). The fully Eurasian-origin sequences were sampled between June and August 2022 from PEI, NL, NS, and NB, while the reassortants were identified in PEI and QC within a week of each other in early June 2022. Detailed analysis of these sequences is the focus of a separate study, and beyond the focus of this paper.
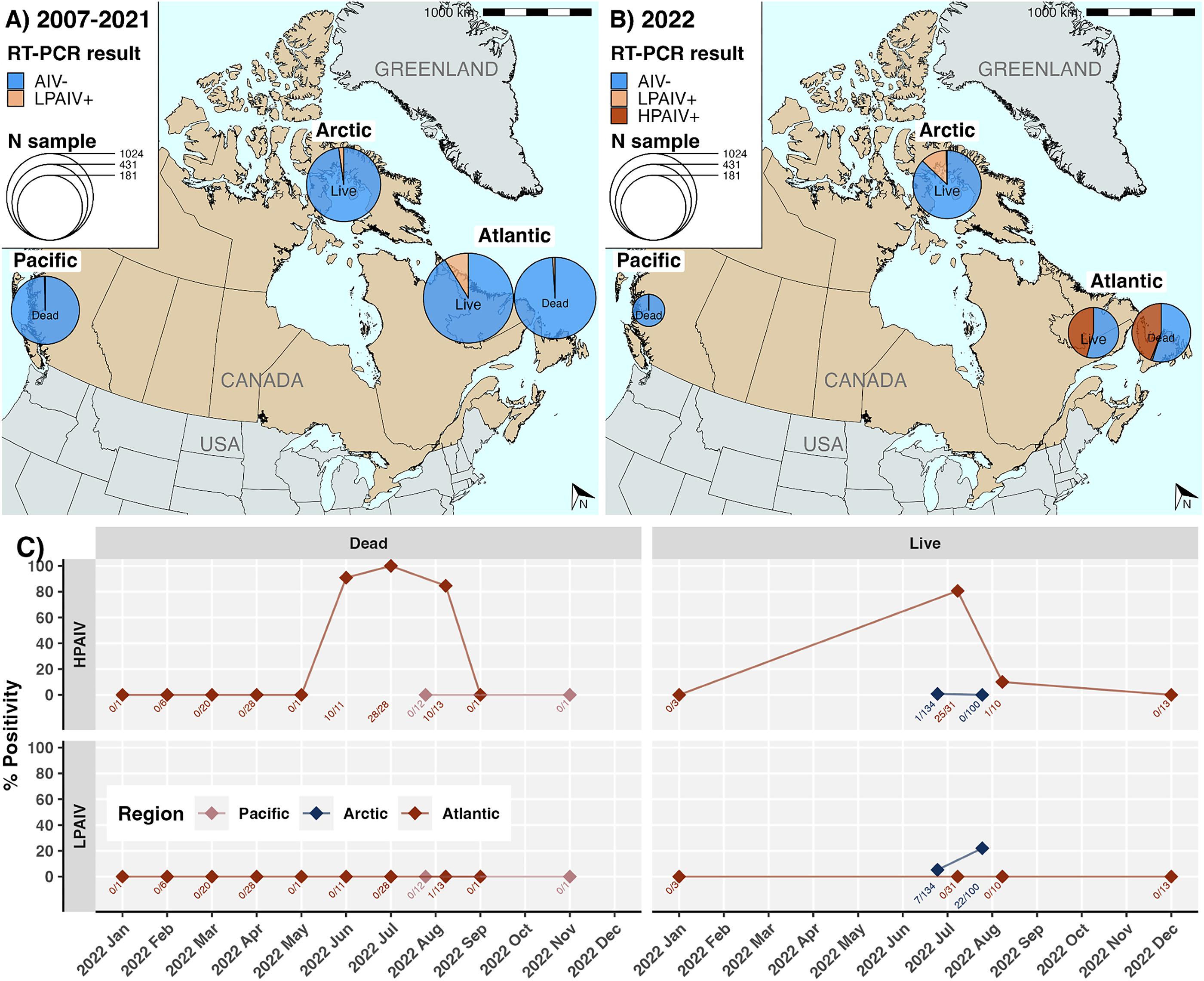
HPAIV was widely detected in murres in 2022 within the Atlantic region: in 48 of 52 dead birds (92.3%) sampled between June and August 2022, and 25 of 31 live breeding birds (80.6%) from July, all most likely locally-breeding COMU (Figs. 2, and 3). HPAIV was additionally detected in the Arctic in one apparently healthy live breeding bird out of 134 TBMU sampled in June. All the AIV detected in NL was HPAIV. However, LPAIV was detected in 43 of 248 live murres (17.3%) sampled in the Arctic in July and August, and in a single dead bird from NB in August. No LPAIV or HPAIV was detected in murres from the Pacific in 2022 (n = 13).
Fig. 2.
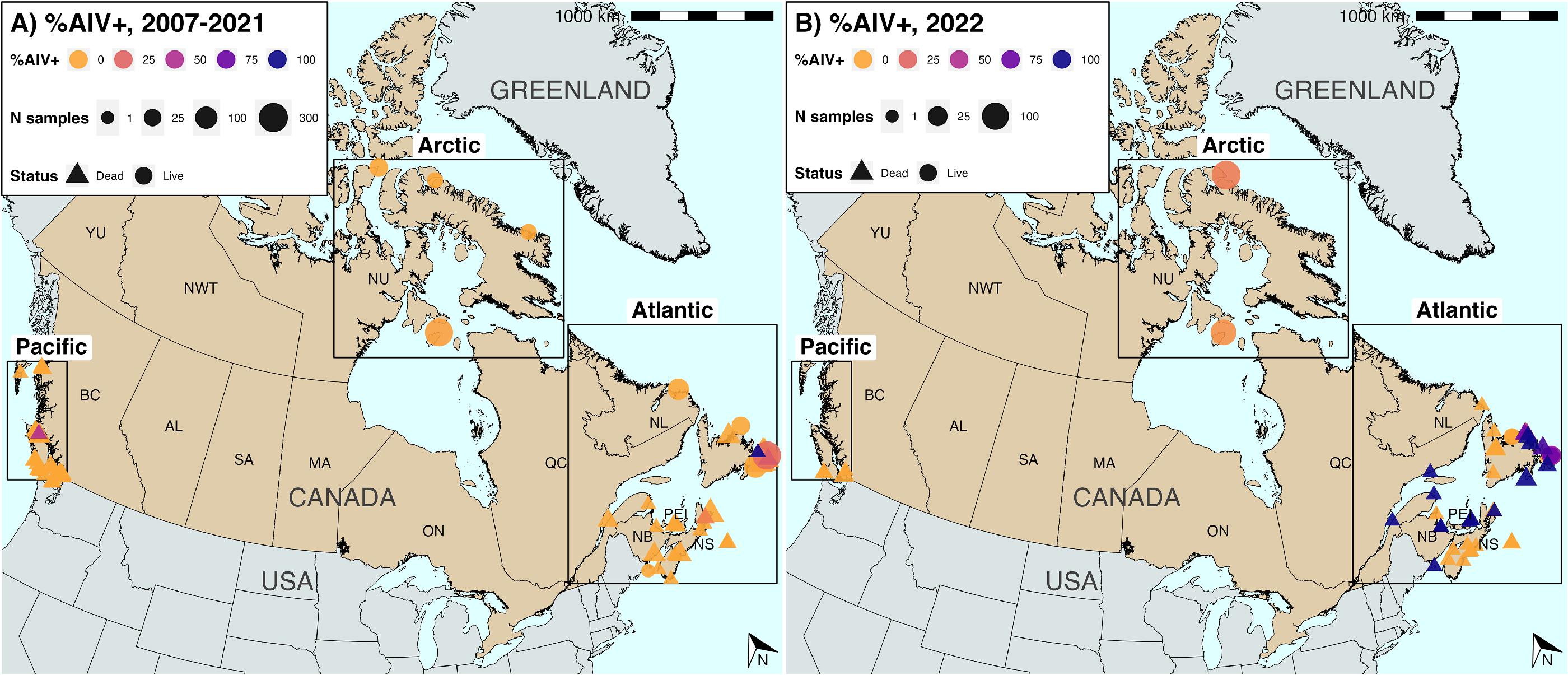
Fig. 3.
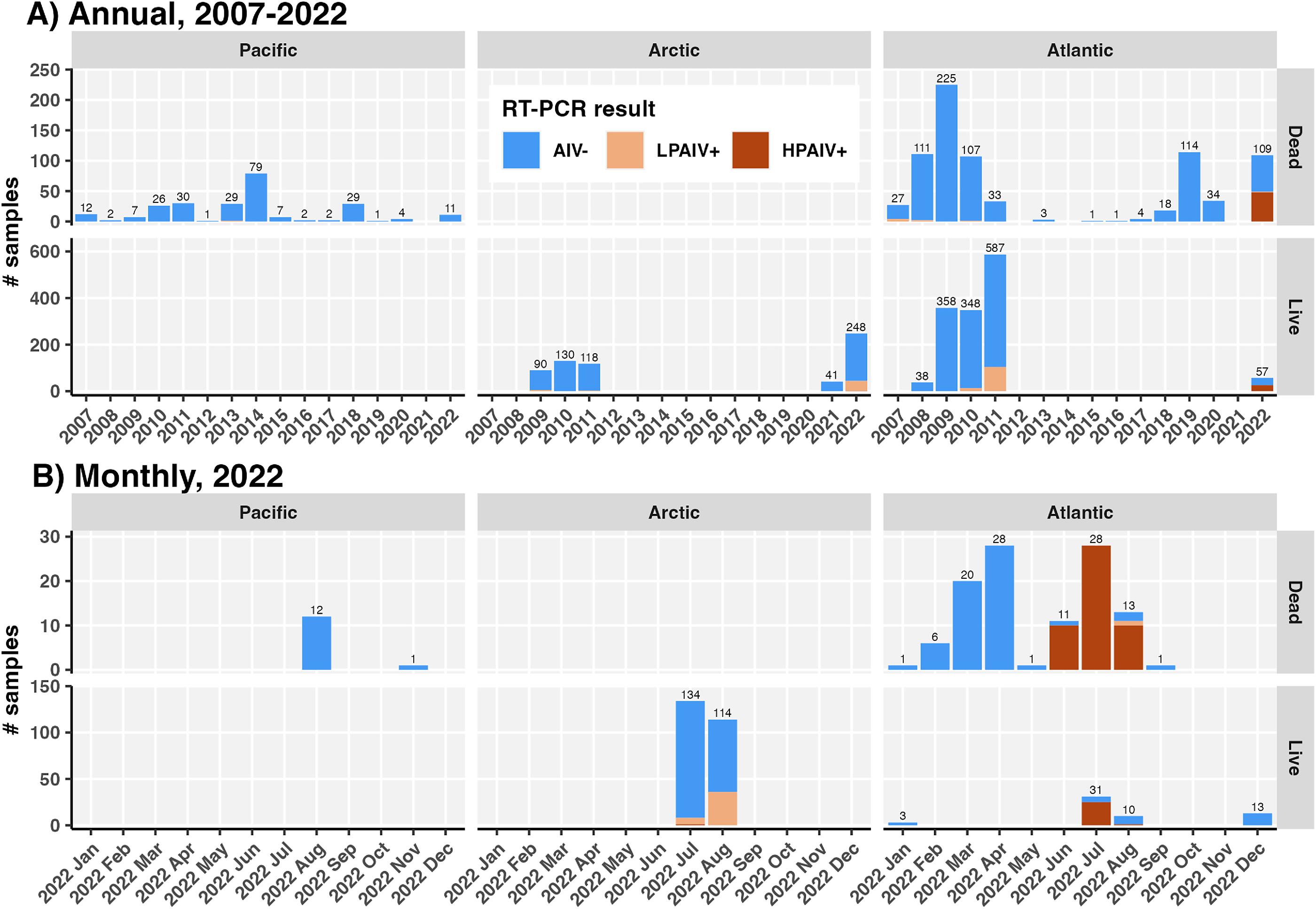
Fig. 4.
Overall, 25.4% (95% CI: 21.4%–30.0%) of all murres sampled in Canada in 2022 were AIV+, of which 59.5% (50.4%–68.1%) were HPAIV+, such that 18.2% (14.6%–22.3%) of all the murres sampled in 2022 were HPAIV+. AIV prevalence was significantly lower in live/harvested murres relative to dead murres, with 23.0% (18.4%–28.2%) AIV+ in live birds compared to 40.2% (31.5%–49.4%) AIV+ in dead birds (χ2, p < 0.001; Table S6). Similarly, HPAIV prevalence was significantly lower in live compared to dead birds, with 8.9% (6.0%–12.8%) HPAIV+ in live birds compared to 39.3% (30.7%–48.6%) HPAIV in dead birds (p < 0.0001). By contrast, LPAIV positivity was significantly lower among dead (0%, 0%–3.8%) than live (16.7%, 12.8%–21.5%) murres (p < 0.001) in 2022. Comparisons of results from live and dead birds should be interpreted cautiously as the groups represent different proportions of species, regions, and months, with different reporting and detection biases (Table 1). In NL, live and dead murres did not have significantly different % AIV+, % LPAIV+, or % HPAIV+ (Fisher’s exact test, all p = 1).
Regional and temporal differences in HPAIV and LPAIV prevalence
There were significant regional variations in % AIV+ (Fisher’s test, p < 0.0001), % HPAIV+ (p < 0.0001), and % LPAIV+ (p < 0.0001) in 2022. Focusing on live birds, AIV prevalence was significantly higher in the Atlantic (45.6%, 32.6%–59.2%) than the Arctic (17.5%, 12.3%–24.3%; p < 0.0001), as was HPAIV prevalence (Atlantic: 45.6%, 32.6%–59.2%; Arctic (0.6%, 0%–3.7%; p < 0.0001). However, it should be noted that live bird collection in the Atlantic is a combination of samples obtained from local breeding COMU and harvested overwintering birds, which includes TBMU breeding in the Arctic that migrate south after breeding (Frederikson et al. 2016). LPAIV prevalence was significantly higher in the Arctic (17%, 11.8%–23.6%) than in the Atlantic (0%, 0%–7.9%; p = 0.002) in 2022. Although LPAIV was not detected in live birds in the Atlantic in 2022, there was one LPAIV+ sample from a dead COMU in NB. No LPAIV or HPAIV was detected in the Pacific region in 2022 among 13 murres found dead. There were differences in percentage positivity across sampling sites within regions (Fig. 2; Figs. S7–S9). Pairwise differences in site-level AIV positivity were significantly lower within-regions (median = 14.6) than between-regions (median = 22.2) (Kruskal–Wallis, p < 0.0001) (Fig. S10).
When the comparison was restricted to live birds sampled in the summer breeding season (June to August), differences between Atlantic and Arctic AIV and HPAIV prevalence were even more pronounced (Table S6) on account of significantly higher AIV and HPAIV prevalence in live birds in the Atlantic in the summer (63.4%, 46.9%–77.4%) compared to the rest of the year (0%, 0%–24.1%; Fisher’s p < 0.0001). This trend was recapitulated in murres found dead in the Atlantic (summer: 93.9% HPAIV+, 82.1%–98.4%; rest of year: 0%, 0%–7.9%; p < 0.0001); outside of summer, no HPAIV or LPAIV was detected at any Atlantic sampling sites in 2022 (Fig. 2). Within the summer breeding season, we compared prevalence in July and August for live birds in the Atlantic and the Arctic separately. AIV prevalence in live birds sampled in the Arctic was significantly higher in August (31.6%, 23.4%–41.0%) compared to July (6%, 2.8%–11.8%, p < 0.0001); however, this may have reflected differences between breeding colonies sampled in these months. In the Atlantic, HPAIV prevalence was significantly higher in July (80.6%, 61.9%–91.9%) compared to August (10%, 0.5%–45.9%) (Fisher’s, p < 0.001).
In a multiple binomial model of AIV positivity in live birds in 2022 adjusted for season, murres sampled in the Atlantic had 8.0 (4.0–16.8) times higher adjusted odds of being AIV+ than murres sampled in the Arctic. The odds of HPAIV positivity in live birds in the Atlantic in 2022 were 428 (82–7906) times higher than in the Arctic, adjusted for season. However, confidence intervals were wide and this relationship should be re-evaluated with longitudinal data.
Long term trends in HPAIV and LPAIV prevalence in murres sampled in Canada
We compared the murre AIV prevalence in 2022 to historical data from 2007–2021, a 15-year period during which 1710 live and 909 dead birds were sampled and tested for AIV, with sampling concentrated primarily pre-2015 (Table 2). Across 2007–2021, 5.1% (4.3%–6%) of 1710 live birds were LPAIV+ compared to 0% (0%–3.7%) of 909 dead birds, although dead birds included Pacific and not Arctic birds, and live did not include any from the Pacific. H5 was not detected in any of 8 AIV+ samples from dead birds and 125 AIV+ from live birds from August 2007 to October 2013. From October 2013–October 2020, none of 325 found dead birds were AIV+. No dead birds were sampled in 2021. Live birds were not sampled for 10 years until July 2021, when none of the 41 live birds sampled in the Arctic were AIV+.
Table 2.
| Category | Group | Live/harvested, N (%) | Dead, N (%) |
|---|---|---|---|
| Total | 1710 (100.0) | 909 (100.0) | |
| Species | COMU | 1215 (71.1) | 411 (45.2) |
| TBMU | 495 (28.9) | 498 (54.8) | |
| Age group | HY | 247 (15.2) | 118 (13.6) |
| AHY | 1382 (84.8) | 747 (86.4) | |
| Unknown | 5 | 44 | |
| Sex | Female | 11 (25.6) | 359 (56.0) |
| Male | 32 (74.4) | 282 (44.0) | |
| Unknown | 1667 | 268 | |
| Region | Pacific | – | 231 (25.4) |
| Arctic | 379 (22.2) | – | |
| Atlantic | 1331 (77.8) | 678 (74.6) | |
| Year | 2007 | – | 39 (4.3) |
| 2008 | 38 (2.2) | 113 (12.4) | |
| 2009 | 448 (26.2) | 232 (25.5) | |
| 2010 | 478 (28.0) | 133 (14.6) | |
| 2011 | 705 (41.2) | 63 (6.9) | |
| 2012 | – | 1 (0.1) | |
| 2013 | – | 32 (3.5) | |
| 2014 | – | 79 (8.7) | |
| 2015 | – | 8 (0.9) | |
| 2016 | – | 3 (0.3) | |
| 2017 | – | 6 (0.7) | |
| 2018 | – | 47 (5.2) | |
| 2019 | – | 115 (12.7) | |
| 2020 | – | 38 (4.2) | |
| 2021 | 41 (2.4) | – |
Note: Percentages by category were calculated excluding missing data. COMU, common murre; TBMU, thick-billed murre; HY, hatch-year; AHY, after hatch-year.
Across regions and seasons, LPAIV prevalence mostly remained below 5%, except for murres found dead in the Atlantic in summer 2007 (n = 20, 20%, 6.6%–44.3%), as well as live murres in June 2009 in the Arctic (n = 62, 8.1%, 3%–18.5%) and July–August 2011 in the Atlantic region (n = 369, 28.2%, 23.7%–33.1%) (Figs. S4–S6). In the Pacific, LPAIV has only been detected once, in a murre found dead in October 2013 (n = 29, 3.4% (0.2%–19.6%)).
We cannot exclude the possibility that HPAIV was present but went undetected during this time, particularly in 2021 during which no dead birds were tested and live data were exclusively from the Arctic, however there is an absence of evidence. Three samples were H5 non-negative in 2007–2009, but these were subsequently found to be LPAIV H5 subtypes of North American origin. Relative to 2011, the last year prior to 2022 with live bird data from the Atlantic, overall AIV prevalence in live murres in the Atlantic region increased significantly from 17.7% (14.8%–21.1%) to 45.6% (32.6%–59.2%) in 2022 (p < 0.0001). Within that, HPAIV prevalence in the Atlantic increased from 0% (0%–3.9%) in 2011 to 45.6% (32.6%–59.2%) in 2022 (p < 0.0001). Correspondingly, LPAIV prevalence in the Atlantic decreased from 17.7% (14.8%–21.1%) in 2011 to 0% (0%–7.9%) in 2022. In the Arctic, overall AIV prevalence in 2022 (17.7%, 13.3%–23.2%) had increased significantly since 2021 (0%, 0%–10.7%; p < 0.01) and 2011 (2.5%, 0.7%–7.8%; p < 0.001), as did LPAIV prevalence (p < 0.01; p < 0.0001, respectively). Although HPAIV prevalence in the Arctic in 2022 (0.4%, 0%–2.6%) was slightly higher than 2021 (0%, 0%–10.7%) or 2011 (0%, 0%–3.9%), we were underpowered to detect significance. All other reported comparisons were sufficiently powered (Table S6).
Sample sizes for future AIV monitoring
Power analyses were used to estimate sample sizes needed in the future for accurate monitoring of changes over time, assuming moderate changes in disease prevalence relative to 2022 (Table S7). In the primary reported chi-squared analysis, an alpha significance level of 0.05, 25% additive change (a moderate effect size), and 80% power were applied; the effects of these parameters on sample sizes were explored (Fig. S11).
For HPAIV (same as for AIV) in NL, where 2022 prevalence was 45.6% (32.6%–59.2%) in 57 samples, in 2023, N = 63 (42–65) samples would be required to detect this moderate effect size with 80% power and α = 0.05, or 201 (99–220) with α = 0.01 (Table S7). Comparing HPAIV in the summer in NL, prevalence was 63.4% (46.9%–77.4%) in 49 samples, N = 38 (10–74) samples are required in summer 2023 to detect a 25% increase with 80% power and α = 0.05, or N = 92 (16–428) at α = 0.01.
For AIV in the Arctic, where 2022 prevalence (all summer samples) was 17.7% (13.3%–23.2%) across 248 samples, 28 (25–32) samples would be required to detect a 25% increase of moderate effect size, with 80% power and α = 0.05, or N = 45 (39–50) at α = 0.01 (Table S7). To detect a smaller increase in AIV in the Arctic of 15% (Table S8), N = 87 (72–104) samples would be required at α = 0.05 and 80% power. For HPAIV in the Arctic, where 2022 summer prevalence was 0.4% (0%–2.6%), to detect a 25% increase, N = 9 (7–14) samples would be required at α = 0.05, or N = 14 (11–21) at α = 0.01. To detect a 15% increase in HPAIV, N = 18 (13–30) samples would be required at α = 0.05, or N = 28 (20–47) at α = 0.01. Where evidence suggests prevalence differs across sites within regions but is similar across months in the summer breeding season, we conservatively recommend approximately 100 samples be tested seasonally at each site, where resources are available and monitoring systems more well-established, for accurate estimation of small to moderate changes in AIV and HPAIV prevalence in murres with an α between 0.01 and 0.05. Collecting samples at multiple time points within the breeding season from the same sites would increase the probability of detecting rapidly transmitted pathogen outbreaks and provide more accurate estimates of average period prevalence.
Discussion
HPAIV H5 clade 2.3.4.4b viruses were widely detected in Canada in 2022 in live and dead murres sampled in the Atlantic and a single live murre from the southeastern Arctic. AIV and HPAIV prevalence in live birds were significantly higher in the Atlantic than Arctic, and were significantly higher during summer months than the rest of the year. HPAIV has not yet been detected in murres in the Pacific, but sampling has been sparse. In comparison with 2007–2021, a period during which HPAIV was not detected in murres, HPAIV and AIV prevalence were significantly elevated in 2022 in the Atlantic and the Arctic.
HPAIV prevalence was low in the Arctic in 2022, but LPAIV prevalence had increased. It is unclear to what extent the relatively higher frequency of LPAIV versus HPAIV in the Arctic is affected by sampling gaps, geographic isolation from affected flyways (lack of opportunity), or other ecological factors. Sampling efforts for AIV monitoring in murres from 2007 to 2022 were highly variable and not comprehensive across breeding colonies, therefore it is possible that HPAIV circulated below detection levels in 2021 in particular; however, there was also a lack of detected excess morbidity or mortality at breeding colonies in 2021. The absence of HPAIV detections in Arctic murres in 2021 corroborates a phylogeographic study that found northern seabird colonies were not implicated among wild birds importing HPAIV in 2021 from Europe to North America via the Atlantic (Caliendo et al. 2022). The patterns of AIV geographic diffusion and interspecies transmission must also take into consideration both virus and host diversity, host behavior, summer temperatures, and geographic distance (Hicks et al. 2022).
We recommend that in future monitoring years 100 samples be collected per site where possible, or per province, to be sufficiently powered to detect moderate increases in HPAIV or AIV prevalence. Additionally, sampling at the same time of year and at the same breeding colonies over time would reduce spatial and temporal (seasonal) bias in sampling, thereby providing more informative data for long term trends assessment. We also recommend recording population parameters, especially annual survival (Frederiksen et al. 2021), alongside disease monitoring to provide more robust information for potential population-level effects.
All the H5 sequences from murres in 2022 fell within clade 2.3.4.4b, and this clade accounted for nearly all the AIV detected in the Atlantic, which is not unexpected based on its preponderance in Canada and the United States (US) since 2021. H5 clade 2.3.4.4b was first detected in Canada in November 2021 in a great black-backed gull (Larus marinus) in St. John’s, NL and then on an exhibition farm amid mass mortality in December 2021 (Caliendo et al. 2022). Phylogenetic analysis of samples from a bald eagle (Haliaeetus leucocephalus) and broiler chickens in BC from February and April 2022 suggested H5 2.3.4.4b was likely first introduced to North America in 2021 via migratory birds through both the Pacific and North Atlantic flyways (Alkie et al. 2022). Prior to the 2021 2.3.4.4b sub-epidemic, high pathogenicity H5 had not been detected since 2015, when H5 clade 2.3.4.4 was identified in BC within small flock chickens and a Cooper’s hawk (Accipiter cooperii; Berhane et al. 2016). From 2014–2015 in the Pacific region of the US, clade 2.3.4.4 (originated from H5N8 then reassorted into H5N1, H5N2) was detected in 2.4% of hunter-harvested birds (Bevins et al. 2016). By June 2015, H5 2.3.4.4 had disappeared in poultry due to interventions such as culling and quarantine, and enigmatically, the clade also appeared to have disappeared in waterfowl (Krauss et al. 2016). However, Ramey et al. responded that this constituted weak evidence of disappearance since the majority of negative samples were collected before the outbreak, in states and provinces where large outbreaks were not reported, and from seabirds, shorebirds, and gulls, as opposed to waterfowl from the order Anseriformes (Ramey et al. 2016); therefore, it is unclear whether H5 2.3.4.4 circulated below detection levels or outside of well-sampled areas, or was indeed eradicated in North America after 2015. Possible mechanisms for this apparent disappearance include existing immunity, antigenic determinants in wild aquatic bird populations, and the cyclic nature of subtype dominance (Krauss et al. 2016). It remains to be seen whether accrual of population-level immunity to H5 2.3.4.4b in wild birds will result in a shift to another subtype or clade, or whether something intrinsic to this clade permits long-term establishment in the wild bird reservoir.
Sampling bias and observational gaps limit the generalizability of our findings. The limitations of small sample sizes and the occurrence of rare events have been considered regularly in ecology (Dixon et al. 2005; Sequeira et al. 2019). When pathogens circulate in wildlife at low prevalence, sample size can influence case ascertainment. The idea that “if you look hard enough you will detect something” (e.g., Mallory et al. 2004) is important to consider when evaluating how prevalence may change over time. While AIV monitoring in murres has not been consistent across the study period, sampling in 2022 did not greatly outpace 2021 in the Atlantic and Arctic; approximately 100 murres in NU were tested annually for AIV prior to 2022. Spurred on by the global concerns around HPAIV, murres were tested in the summer of 2021 as part of an anticipatory effort to increase surveillance (Fig. 3A). Given that sampling effort has not significantly changed, but we see differences in AIV and HPAIV prevalence, this suggests that these are likely reflecting population-level changes. Other sampling biases that may have affected the results are ease of access for live and dead birds, spatially clustered sampling, survival bias among live birds, and pathogenicity bias in birds found dead.
By aggregating data into regions, this analysis was subject to bias due to the modifiable aerial unit problem, whereby summaries are affected by the size and shape of the aggregation unit (Dark and Bram, 2007). Pairwise site-level differences in AIV prevalence were significantly lower within- than between-region, suggesting that regional delineations reflected to some extent meaningful ecological, migratory, and geographic groupings. In a related point, we detected differences in AIV prevalence between breeding colonies and further sampling is required over time to understand if this is a reproducible occurrence. This is consistent with the spatiotemporal variation in AIV prevalence that has been described in wild migratory birds of North America (Munster et al. 2007).
There is an increased need to have up-to-date estimates of prevalence, disease burden, and mortality attributable to HPAIV to inform seabird management and public health risk assessments. At the population level, monitoring viral prevalence and disease burden to determine background and AIV-associated mortality is critical. Disease burden estimates can be incorporated into population status models that consider multiple sources of mortality, where mortality exceeding a threshold can be used as a criterion to warrant taking action with wildlife management tools such as harvest limits and education (Fig. 1B).
Gaps in viral monitoring make it challenging to detect outbreaks and understand whether they are increasing in frequency or severity. Improving viral monitoring systems for data collection, curation, and analysis in wildlife will improve our ability to rapidly detect, respond to, and learn from outbreaks (Verhagen et al. 2021; USGS 2022; WHISPers 2023). There is a critical need to develop evidence-based, wildlife viral monitoring tools by collaborating across agencies to analyze existing data, design programs that are cost-effective, and produce high quality data (Stephen et al. 2004; Giacinti et al. 2022). In Canada, some of the challenges faced by wildlife health surveillance programs include a lack of a regulatory framework surrounding cooperation and coordination across government levels and agencies, as well as complex sampling biases and competing priorities (Giacinti et al. 2022). This results in waves of interest, and the monitoring taking place is resource-dependent and often triggered by some event, such as mass mortalities in wild birds or domestic culling, accompanied by economic losses and public impetus. Resources wane with weakening public interest and eventually are diverted to the next emerging issue. This can be seen in the patterns of AIV monitoring in murres in Canada as described above.
Reactive outbreak mobilization within wildlife viral monitoring programs has also been exemplified for coronaviruses (Ntumvi et al. 2022; Ruiz-Avarena et al. 2022), paramyxoviruses (Drexler et al. 2012), Nipah virus (Epstein et al. 2020), Zika virus (Metsky et al. 2017), Ebola virus (Leroy et al. 2004), and influenza viruses (Bevins et al. 2014; Alkie et al. 2022). There are exceptions to this globally, such as for West Nile virus (Kilpatrick 2011). Although Canada is widely seen to have quite robust wildlife health monitoring programs, this is largely driven by the dedication of regional personnel, and there are clear deficiencies for an ongoing monitoring program. Again, this is demonstrated in the murre dataset as regions differed in their ability to mobilize to increase AIV surveillance at different levels.
From a murre management and conservation perspective, combining viral monitoring with other data, such as presence of other microbes, contaminants, and immune function (Mallory et al. 2010), that affect birds’ survival can reveal cumulative and pleiotropic effects on acute survival and reproduction. In birds, for example, higher prevalence of parasites has been linked to reduced reproductive fecundity (Hudson and Dobson, 1991), and high contaminant levels of mercury may be correlated with more frequently severe viral infections (Teitelbaum et al. 2022). Human activities resulting in environmental and climate changes, as well as habitat loss could alter viral persistence in the environment and provide additional opportunities for viral reassortment and cross-species transmission (Morin et al. 2018; Carlson et al. 2022). Pathogen monitoring is only one aspect of a more comprehensive understanding of wild birds’ population dynamics.
Murre populations experience mortality for a variety of reasons outside of HPAIV, including as bycatch in fisheries gear, oil spill events, poor nutrition, extreme weather, and harvest (Gaston et al. 2002; Wiese et al. 2004; Wiese and Robertson 2004; Clairbaux et al. 2019; Frederiksen et al. 2019). Although tracking banded individuals and birds with logging devices has clarified murre wintering strategies and site fidelity (McFarlane Tranquilla et al. 2014), as well as size and composition of wintering populations (Frederiksen et al. 2016), tracking survival of birds across their range could add a finer resolution of when, where, and possibly how birds are dying. Serological samples paired with swabs from live birds would clarify the role of prior exposure and immunity towards estimated incidence and survival rates. Continued monitoring for HPAIV in birds found dead, including beached bird surveillance, is an additional important surveillance tool to quantify changes in HPAIV diversity and associated mortality.
The population-level impacts of HPAIV are relevant for the sustainability and availability of murres as a food source for subsistence and sociocultural value. The results presented here suggest that during this harvest season, typically 3.5 months between September to March in Atlantic Canada, murres experience low to negligible levels of active infections. The hunter health risk of AIV is low if harvested birds are handled and cooked properly (Canadian Food Inspection Agency 2022). Nonetheless, this information is important to be able to share with harvesters as it relates to bird handling and food security questions. Thus, even though AIV prevalence during the winter murre harvest is likely low, continued winter screening of murres to inform public health advice, and detect any changes in winter prevalence of AIV is advisable, again at the same sample size targets (∼100 individuals per region).
With continuous measurements and predefined action plans, HPAIV monitoring could evolve into HPAIV surveillance (Hoinville et al. 2013). There are often no predefined conservation and management policies or actions for surveillance, but they can include adjusting hunter permitting and safety guidance, education campaigns to reduce exposure, or wildlife vaccination for targeted species of conservation concern, based upon estimates of disease prevalence to counterbalance excess bird mortality (Fig. 1). For the case of murres in Canada, surveillance of AIV is important for both tracking the virus and pathology in murre populations as part of a conservation program, and to inform public health discussions around hunter safety and guidelines related to human ingestion. Within a One Health monitoring framework, which emphasizes the connections between the health of humans, wild and domestic animals, and environments, interdisciplinary collaboration is key to understand, control and prevent zoonoses, and more broadly to enhance ecosystem health (Gibbs 2014).
Conclusions
This analysis of spatiotemporal trends in the prevalence of AIV in murres across Canada serves as a case study of wildlife viral epidemiological monitoring, which could be generalized more broadly for additional species. We are best equipped to understand and mitigate the cumulative impacts of pathogenic threats by quantifying the microbial burden on bird health and survival in the context of the complexity of other health-determining factors.
Acknowledgements
We gratefully acknowledge all the agencies, laboratories, field crews, individuals, communities, hunters, and laboratory staff who participated in sample collection, transport, molecular analyses, and data curation. Special thanks to Trevor Thompson and Cynthia Pekarik for supporting and enabling this work. We also thank the British Columbia Ministry of Agriculture and Food Animal Health Centre staff and pathologists, wildlife rehabilitation centres, Birds Studies Canada, Fisheries and Oceans Canada, Ecotrust Canada, and Parks Canada. Funding for this study was provided by Environment and Climate Change Canada. We thank the referees for providing comments on our manuscript.
References
Alkie T.N., Cox S., Embury-Hyatt C., Stevens B., Pople N., Pybus M.J., et al. 2023. Characterization of neurotropic HPAI H5N1 viruses with novel genome constellations and mammalian adaptive mutations in free-living mesocarnivores in Canada. Emerging Microbes & Infections, 12(1): 2186608.
Alkie T.N., Lopes S., Hisanaga T., Xu W., Suderman M., Koziuk J., et al. 2022. A threat from both sides: multiple introductions of genetically distinct H5 HPAI viruses into Canada via both East Asia-Australasia/Pacific and Atlantic flyways. Virus Evolution, 8(2): veac077.
Banyard A.C., Lean F.Z.X., Robinson C., Howie F., Tyler G., Nisbet C., et al. 2022. Detection of highly pathogenic avian influenza virus H5N1 Clade 2.3.4.4b in great skuas: a species of conservation concern in Great Britain. Viruses, 14(2): 212.
Berhane Y., Kobasa D., Embury-Hyatt C., Pickering B., Babiuk S., Joseph T., et al. 2016. Pathobiological characterization of a novel reassortant highly pathogenic H5N1 virus isolated in British Columbia, Canada, 2015. Scientific Reports, 6(1): 23380.
Bevins S.N., Dusek R.J., White C.L., Gidlewski T., Bodenstein B., Mansfield K.G., et al. 2016. Widespread detection of highly pathogenic H5 influenza viruses in wild birds from the Pacific Flyway of the United States. Scientific Reports, 6: 1–9.
Bevins S.N., Pedersen K., Lutman M.W., Baroch J.A., Schmit B.S., Kohler D., et al. 2014. Large-scale avian influenza monitoring in wild birds throughout the United States. PLoS One, 9(8): e104360–8.
Caliendo V., Lewis N.S., Pohlmann A., Baillie S.R., Banyard A.C., Beer M., et al. 2022. Transatlantic spread of highly pathogenic avian influenza H5N1 by wild birds from Europe to North America in 2021. Scientific Reports, 12(1): 11729.
Canadian Food Inspection Agency. 2022. Status of ongoing avian influenza response by province. Available from https://inspection.canada.ca/animal-health/terrestrial-animals/diseases/reportable/avian-influenza/hpai-in-canada/status-of-ongoing-avian-influenza-response/eng/1640207916497/1640207916934 [accessed 28 April 2023].
Canadian Wildlife Health Cooperative. 2022. Inter-agency Wild Bird Avian Influenza Survey update—December 31, 2022. Available from http://www.cwhc-rcsf.ca/avian_influenza_biweekly_reports.php.
Carlson C.J., Albery G.F., Merow C., Trisos C.H., Zipfel C.M., Eskew E.A., et al. 2022. Climate change increases cross-species viral transmission risk. Nature, 607(7919): 555–562.
Clairbaux M., Fort J., Mathewson P., Porter P., Strøm H., Grémillet D. 2019. Climate change could overturn bird migration: Transarctic flights and high-latitude residency in a sea ice free Arctic. Sci Rep, 9: 17767.
Dark S.J., Bram D. 2007. The modifiable areal unit problem (MAUP) in physical geography. Progress in Physical Geography: Earth and Environment, 31: 471–479.
Dixon P.M., Ellison A.M., Gotelli N.J. 2005. Improving the precision of estimates of the frequency of rare events. Ecology, 86(5): 1114–1123. Available from https://www.jstor.org/stable/3450872.
Drexler J.F., Corman V.M., Müller M.A., Maganga G.D., Vallo P., Binger T., et al. 2012. Bats host major mammalian paramyxoviruses. Nature Communications, 3(1): 796.
Epstein J.H., Anthony S.J., Islam A., Kilpatrick A.M, Ali Khan S., Balkey M.D., et al. 2020. Nipah virus dynamics in bats and implications for spillover to humans. Proceedings of the National Academy of Sciences, 117: 29190.
Falchieri M., Reid S.M., Ross C.S., James J., Byrne A.M., Zamfir M., et al. 2022. Shift in HPAI infection dynamics causes significant losses in seabird populations across Great Britain. Veterinary Record, 191: 294–296.
Frederiksen M., Descamps S., Elliott K.H., Gaston A.J., Huffeldt N.P., Kolbeinsson Y., et al. 2021. Spatial variation in vital rates and population growth of thick-billed murres in the Atlantic Arctic. Marine Ecology Progress Series, 672:1–13.
Frederiksen M., Descamps S., Erikstad K.E., Gaston AJ., Gilchrist H.G, Grémillet D., et al. 2016. Migration and wintering of a declining seabird, the thick-billed murre Uria lomvia, on an ocean basin scale: conservation implications. Biological Conservation, 200: 26–35.
Frederiksen M., Linnebjerg J.F., Merkel F.R., Wilhelm S.I., Robertson G.J. 2019. Quantifying the relative impact of hunting and oiling on Brünnich's guillemots in the north-west Atlantic. Polar Research, 38.
gadm.org. 2022. gadm v4.1 - Greenland. Available from https://gadm.org/download_country.html.
Gamarra-Toledo V., Plaza P.I., Inga G., Gutiérrez R., García-Tello O., Valdivia-Ramírez L., et al. 2023. First mass mortality of marine mammals caused by highly pathogenic influenza virus (H5N1) in South America. [preprint: bioRxiv]. bioRxiv.
Gaston A.J., Hipfner J.M, Campbell D. 2002. Heat and mosquitoes cause breeding failures and adult mortality in an Arctic-nesting seabird. IBIS, 144(2): 185–191.
Giacinti J.A., Jane Parmley E., Reist M., Bayley D., Pearl D.L., Jardine C.M. 2022. Canadian wildlife health surveillance—patterns, challenges and opportunities identified by a scoping review. FACETS, 7(1): 25–44.
Giacinti J.A., Signore A.V., Jones M.E.B., Bourque L., Lair S., Jardine C., et al. 2023. Avian influenza viruses in wild birds in Canada following incursions of highly pathogenic H5N1 virus from Eurasia in 2021/2022. [preprint: bioRxiv]. bioRxiv.
Gibbs E.P.J. 2014. The evolution of One Health: a decade of progress and challenges for the future. Veterinary Record, 174(4): 85.
Granter A., Wille M., Whitney H., Robertson G.J., Ojkic D., Lang A.S. 2010. The genome sequence of an H11N2 avian influenza virus from a thick-billed murre (Uria lomvia) shows marine-specific and regional patterns of relationships to other viruses. Virus Genes, 41: 224–230.
Harvey J.A., Mullinax J.M., Runge M.C., Prosser D.J. 2023. The changing dynamics of highly pathogenic avian influenza H5N1: next steps for management & science in North America. Biological Conservation, 282: 110041.
Hicks J.T., Edwards K., Qiu X., Kim Do-K, Hixson J.E., Krauss S., et al. 2022. Host diversity and behavior determine patterns of interspecies transmission and geographic diffusion of avian influenza A subtypes among North American wild reservoir species. PLoS Pathogens, 18(4): e1009973.
Hoinville L.J., Alban L., Drewe J.A., Gibbens J.C., Gustafson L., Häsler B., et al. 2013. Proposed terms and concepts for describing and evaluating animal-health surveillance systems. Prev Vet Med., 1;112(1-2): 1–12.
Huang Y., Robertson G.J., Ojkic D., Whitney H., Lang AS. 2014. Diverse inter-continental and host lineage reassortant avian influenza A viruses in pelagic seabirds. Infection, Genetics and Evolution, 22: 103–111.
Hudson, P.J., Dobson, A.P. 1991. Control of parasites in natural populations: nematode and virus infections of Red Grouse. In C M Perrins, J -D Lebreton, and G J M Hirons (eds), Bird Population Studies: Relevance To Conservation And Management Oxford Academic.
Kilpatrick A.M. 2011. Globalization, land use, and the invasion of West Nile virus. Science, 334(6054): 323–327.
Krauss S., Stallknecht D.E., Slemons R.D., Bowman A.S., Poulson R.L., Nolting J.M., et al. 2016. The enigma of the apparent disappearance of Eurasian highly pathogenic H5 clade 2.3.4.4 influenza A viruses in North American waterfowl. Proceedings of the National Academy of Sciences, 113(32): 9033–9038.
Lang A.S., Lebarbenchon C., Ramey A.M., Robertson G.J., Waldenström J., Wille M. 2016. Assessing the role of seabirds in the ecology of influenza A viruses. Avian Diseases, 60: 378–386.
Leroy EM., Rouquet P., Formenty P., Souquière S., Kilbourne A., Froment J.-M., et al. 2004. Multiple Ebola virus transmission events and rapid decline of Central African wildlife. Science, 303(5656): 387–390.
Liu J., Xiao H., Lei F., Zhu Q., Qin K., Zhang X.-W., et al. 2005. Highly pathogenic H5N1 influenza virus infection in migratory birds. Science, 309(5738): 1206.
Mallory M.L., Kiff L., Clark R.G., Bowman T., Blums P., Mednis A., Alisauskas R.T. 2004. The occurrence of runt eggs in waterfowl clutches. Journal of Field Ornithology, 75(3): 209–217.
Mallory M.L., Robinson S.A., Hebert C.E., Forbes M.R. 2010. Seabirds as indicators of aquatic ecosystem conditions: a case for gathering multiple proxies of seabird health. Marine Pollution Bulletin, 60(1): 7–12.
Mcfarlane Tranquilla L.A., Montevecchi W.A., Fifield D.A., Hedd A., Gaston A.J., Robertson G.J., Phillips R.A. 2014. Individual winter movement strategies in two species of murre (Uria spp.) in the Northwest Atlantic. PLoS One, 9(4): e90583.
Mchugh M.L. 2013. The chi-square test of independence. Biochemia Medica, 23(2): 143–149.
Metsky H.C., Matranga C.B., Wohl S., Schaffner S.F., Freije C.A., Winnicki S.M., et al. 2017. Zika virus evolution and spread in the Americas. Nature, 546(7658): 411–415.
Morin C.W., Stoner-Duncan B., Winker K., Scotch M., Hess J.J., Meschke JS., et al. 2018. Avian influenza virus ecology and evolution through a climatic lens. Environment International, 119: 241–249.
Munster V.J., Baas C., Lexmond P., Waldenström J., Fransson T., Rimmelzwaan G.F., et al. 2007. Spatial, temporal, and species variation in prevalence of influenza A viruses in wild migratory birds. PLoS Pathog., 11;3(5): e61.
National Oceanographic and Atmospheric Administration (NOAA) Fisheries. 2022. Recent increase in seal deaths in Maine linked to avian flu. Available from https://www.fisheries.noaa.gov/feature-story/recent-increase-seal-deaths-maine-linked-avian-flu [accessed October 2023].
Ntumvi N.F., Ndze V.N., Gillis A., Le Doux Diffo J., Tamoufe U., Takuo J.-M., et al. 2022. Wildlife in Cameroon harbor diverse coronaviruses, including many closely related to human coronavirus 229E. Virus Evolution, 8(1): veab110.
Ramey A.M., Hill N.J., Deliberto T.J., Gibbs S.E.J., Camille Hopkins M., Lang A.S., et al. 2022. Highly pathogenic avian influenza is an emerging disease threat to wild birds in North America. The Journal of Wildlife Management, 86(2).
Ramey A.M., Spackman E., Kim-Torchetti M., Deliberto T.J. 2016. Weak support for disappearance and restricted emergence/persistence of highly pathogenic influenza A in North American waterfowl. Proceedings of the National Academy of Sciences, 113(43): E6551–E6552.
Ruiz-Aravena M., McKee C., Gamble A., Lunn T., Morris A., Snedden C.E., et al. 2022. Ecology, evolution and spillover of coronaviruses from bats. Nat Rev Microbiol., 20: 299–314.
Sequeira A.M.M., Heupel M.R., Lea M.‐A., Eguíluz V.M., Duarte C.M., Meekan M.G., et al. 2019. The importance of sample size in marine megafauna tagging studies. Ecological Applications, 29(6): e01947. Available from https://www.jstor.org/stable/26785698.
Spackman E., Senne D.A., Myers T.J., Bulaga L.L., Garber L.P., Perdue M.L., et al. 2002. Development of a real-time reverse transcriptase PCR assay for type A influenza virus and the avian H5 and H7 hemagglutinin subtypes. Journal of Clinical Microbiology, 40(9): 3256–3260.
Statistics Canada. 2019. 2016 Census—Boundary files. Available from https://www12.statcan.gc.ca/census-recensement/2011/geo/bound-limit/bound-limit-2016-eng.cfm.
Stephen C., Artsob H., Bowie W.R., Drebot M., Fraser E., Leighton T., et al. 2004. Perspectives on emerging zoonotic disease research and capacity building in Canada. Canadian Journal of Infectious Diseases and Medical Microbiology, 15(6): 339–344.
Stephen C., Duff J.P., Gavier-Widen D., Ryser-Degiorgis M.P., Uhart M.M., Sleeman J., et al. 2018. Proposed attributes of national wildlife health programmes. Revue Scientifique et Technique de l'OIE, 37(3): 925–936.
Teitelbaum C.S., Ackerman J.T., Hill M.A., Satter J.M., Casazza M.L., De La Cruz S.E.W., et al. 2022. Avian influenza antibody prevalence increases with mercury contamination in wild waterfowl. Proceedings of the Royal Society B: Biological Sciences, 289(1982): 20221312.
United States Census Bureau. 2019. Cartographic boundary files. Available from https://www.census.gov/geographies/mapping-files/time-series/geo/cartographic-boundary.html.
United States Geological Survey (USGS) National Wildlife Health Center. 2022. Distribution of highly pathogenic avian influenza in North America, 2021/2022. Available from https://www.usgs.gov/centers/nwhc/science/distribution-highly-pathogenic-avian-influenza-north-america-20212022 [accessed 12 May 2023].
Verhagen J.H., Fouchier R.A.M., Lewis N. 2021. Highly pathogenic avian influenza viruses at the wild–domestic bird interface in Europe: future directions for research and surveillance. Viruses., 13(2): 212.
Wiese F.K., Robertson G.J. 2004. Assessing seabird mortality from chronic oil discharges at sea. Journal of Wildlife Management, 68(3): 627–638.
Wiese F.K., Robertson G.J., Gaston A.J. 2004. Impacts of chronic marine oil pollution and the murre hunt in Newfoundland on thick-billed murre Uria lomvia populations in the eastern Canadian Arctic. Biological Conservation, 116(2): 205–216.
Wildlife Health Information Sharing Partnership—event reporting system (WHISPers). 2023. Available from https://whispers.usgs.gov/home [accessed 12 May 2023].
Wille M., Huang Y., Robertson G.J., Ryan P., Wilhelm S.I., Fifield D., et al. 2014. Evaluation of seabirds in Newfoundland and Labrador, Canada, as hosts of influenza A viruses. Journal of Wildlife Diseases, 50(1): 98–103.
Supplementary material
Supplementary Material 1 (DOCX / 4.09 MB).
- Download
- 4.09 MB
Information & Authors
Information
Published In

FACETS
Volume 9 • 2024
Pages: 1 - 13
Editor: Jian Liu
History
Received: 19 October 2023
Accepted: 15 March 2024
Version of record online: 10 July 2024
Copyright
© 2024 Authors Rahman, Wight, Hargan, Lang, Mallory, Elliott, Ojkic, Lair, and Jones, and The Crown. This work is licensed under a Creative Commons Attribution 4.0 International License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Data Availability Statement
Data available upon request.
Key Words
Sections
Subjects
Authors
Author Contributions
Conceptualization: AM, JG, GJR, JP
Data curation: AM, JG, IR, JW, KH, ASL, MM, GJR, KE, DO, SL, MJ, YB, GG, LW, SIW, MGCB, JP
Formal analysis: AM, JG, GJR, JP
Funding acquisition: JG, KH, ASL, MM, GJR, KE, DO, SL, MJ, YB, GG, LW, SIW, MGCB, JP
Investigation: AM, JG, IR, JW, KH, ASL, MM, GJR, KE, DO, SL, MJ, YB, GG, LW, SIW, MGCB, JP
Methodology: AM, JG, JW, ASL, MM, GJR, DO, MGCB, JP
Project administration: JG, GJR, JP
Resources: JG, KH, ASL, MM, GJR, KE, DO, SL, MJ, YB, GG, LW, SIW, MGCB, JP
Software: AM, GJR
Supervision: JG, GJR, JP
Validation: AM, JG, ASL, MM, GJR, KE, DO, SL, MJ, YB, GG, LW, SIW, MGCB, JP
Visualization: AM, GJR
Writing – original draft: AM, JG, GJR, JP
Writing – review & editing: AM, JG, IR, JW, KH, ASL, MM, GJR, KE, DO, SL, MJ, YB, GG, LW, SIW, MGCB, JP
Competing Interests
The authors declare there are no competing interests.
Funding Information
Metrics & Citations
Metrics
Other Metrics
Citations
Cite As
Angela McLaughlin, Jolene Giacinti, Ishraq Rahman, Jordan Wight, Kathryn Hargan, Andrew S. Lang, Mark L. Mallory, Gregory J. Robertson, Kyle Elliot, Davor Ojkic, Stéphane Lair, Megan Jones, Yohannes Berhane, Grant Gilchrist, Laurie Wilson, Sabina I. Wilhelm, Michael G.C. Brown, and Jennifer F. Provencher. 2024. Spatiotemporal patterns of low and highly pathogenic avian influenza virus prevalence in murres in Canada from 2007 to 2022—a case study for wildlife viral monitoring. FACETS.
9: 1-13.
https://doi.org/10.1139/facets-2023-0185
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
Cited by
1. Examining avian influenza virus exposure in seabirds of the northwest Atlantic in 2022 and 2023 via antibodies in eggs