Survey of mercury in boreal chorus frog (Pseudacris maculata) and wood frog (Rana sylvatica) tadpoles from wetland ponds in the Prairie Pothole Region of Canada
Abstract
Tadpoles are important prey items for many aquatic organisms and often represent the largest vertebrate biomass in many fishless wetland ecosystems. Neurotoxic mercury (Hg) can, at elevated levels, decrease growth, lower survival, and cause developmental instability in amphibians. We compared total Hg (THg) body burden and concentration in boreal chorus frog (Pseudacris maculata) and wood frog (Rana sylvatica) tadpoles. Overall, body burden and concentration were lower in boreal chorus frog tadpoles than wood frog tadpoles, as expected, because boreal chorus frog tadpoles consume at lower trophic levels. The variables species, stage, and mass explained 21% of total variation for body burden in our models but had negligible predictive ability for THg concentration. The vast majority of the remaining variation in both body burden and THg concentration was attributable to differences among ponds; tadpoles from ponds in three areas had considerably higher THg body burden and concentration. The pond-to-pond differences were not related to any water chemistry or physical parameter measured, and we assumed that differences in wetland geomorphology likely played an important role in determining Hg levels in tadpoles. This is the first report of Hg in frog tadpoles in the Prairie Pothole Region of North America.
Introduction
The elevated transport and deposition of mercury (Hg) to remote pristine areas, such as the prairie region of North America, occurs via anthropogenic Hg emissions (Schuster et al. 2002). Hg in the atmosphere is deposited onto the landscape via wet or dry deposition (Hall et al. 2005; Graydon et al. 2008; Weiss-Penzias et al. 2016), and once HgII enters aquatic ecosystems it can be readily transformed into the neurotoxic organic methylmercury (MeHg; Gilmour and Henry 1991; Branfireun et al. 1999). Although MeHg can be produced abiotically, most is methylated via microbial activity in wetlands and lakes, where HgII is reduced and bound to a methyl group (Boening 2000; Hintelmann 2010). Once present in aquatic ecosystems, MeHg can be taken up and biomagnified with increasing trophic level (Morel et al. 1998; Rimmer et al. 2005; Scheuhammer et al. 2007; Hall et al. 2009). Effects of MeHg exposure in humans and wildlife include reduced reproductive rates, hormonal and immune system changes, altered brain chemistry, behavioral or visual abnormalities, and mortality (Scheuhammer et al. 2007; Hightower 2009).
The Prairie Pothole Region (PPR) of North America is an area of abundant isolated wetland ponds that are usually small, shallow, and productive water bodies surrounded by riparian zones that are usually dominated by cattails (Typha latifolia), bulrushes (Schoenoplectus spp.), sedges (Carex spp. and Scirpus spp.), and willows (Salix spp.) (Batzer and Sharitz 2006; Johnson et al. 2010; Hayashi et al. 2016). These wetlands provide many ecosystem services including groundwater recharge, flood control, water quality, and carbon and nutrient sequestration (van der Kamp et al. 2003; Hayashi et al. 2016). They are also important breeding and nesting habitats for avian wildlife in North America (Batt et al. 1989). Atmospherically deposited Hg may be readily methylated (Hoggarth et al. 2015) and accumulated (Sando et al. 2007; Hall et al. 2009) in these environments because prairie wetlands have potentially high dissolved organic carbon (DOC) concentration, a warm temperature in the summer months, and low dissolved oxygen; all variables which lead to anoxic conditions in which the main methylating organisms (sulfate and iron reducing bacteria and methanogens (Gilmour et al. 2013; Parks et al. 2013)) thrive (Paranjape and Hall 2017).
Amphibians can be highly sensitive to MeHg because it may reduce reproductive success, growth, and survival, especially in the early developmental stages (Zillioux et al. 1993; Unrine et al. 2004, 2007; Bank et al. 2007b; Bergeron et al. 2010a). Despite the sensitivity of amphibians to Hg, fish are the most studied organism in terms of MeHg toxicity. Within aquatic habitats, amphibians and other mid-level trophic consumers are prey for fish and other aquatic biota, which in turn may be consumed by humans or other top predators of the ecosystem. Thus, lower trophic levels can mediate the transfer of MeHg from aquatic to terrestrial ecosystems (Bank et al. 2007a). Although tadpoles are important prey for many fish and waterfowl (Bergeron et al. 2010b), there is a lack of studies focused on amphibians compared with other aquatic vertebrates (Bradford et al. 2012). Furthermore, tadpole biomass is often the largest of all vertebrates in many fishless wetland ecosystems, making them an important part of ecosystem functioning (Ugarte et al. 2005). Finally, Hg concentration and body burden in frog tadpoles from the PPR have not been reported in the literature. Our objective here was to compare total Hg (THg) concentration in herbivorous boreal chorus frog (Pseudacris maculata Agassiz, 1850) and omnivorous wood frog (Rana sylvatica LeConte, 1825) tadpoles from Saskatchewan wetlands. By studying lower trophic levels, inferences can be made about higher level consumers, and precautions can be implemented if a high concentration of Hg is found. Frog tadpoles are a good biomonitor of processes within wetland habitats as almost all of their food sources are aquatic, unlike other amphibians or adult frogs (Hothem et al. 2010). Global amphibian population declines have become a concern (Stuart et al. 2004) and Hg monitoring in wetland amphibians can be useful as some of these declines may be attributed to environmental Hg contamination (Corn 2000; Ficken and Byrne 2013).
Methods
Site selection
Twenty-two sampling sites were located across central Saskatchewan (Table 1 and Fig. 1). From 1981 to 2010 this region had daily mean temperatures ranging from a mean winter temperature of −14.7 °C in January to a mean summer temperature of 18.9 °C in July (Environment Canada 2016). Over the same period, mean annual precipitation was 389.7 mm, with approximately 26% as snowfall (Environment Canada 2016). The area of the wetland ponds ranged from 0.2 to 13 ha (data not shown), and the maximum depth at the time of sampling was between 0.45 and 3.15 m (Table 1). Ponds did not have surface water connectivity, and although we did not measure groundwater dynamics, prairie wetlands typically have limited surface and groundwater connectivity (Hayashi et al. 2016). Ponds were surrounded by conventional farmland, organic farmland, minimum tillage farmland, or wildlife areas (Donald et al. 2001). Conventional and organic farms were generally tilled three times a year, whereas minimum tillage farms limited tilling to once a year or were not tilled at all (Donald et al. 2001). Wildlife areas were parcels of Saskatchewan grasslands that were not altered for agricultural use. These areas included native prairies, nature park reserves, and recreational park reserves. Ponds were selected in a previous study (Donald et al. 2001) examining the impact of adjacent farming practices on pesticide concentrations. Ponds were named according to family name of a resident of the area and adjacent farming practices. Although we do not consider the impact of farming practices here, we have maintained the names for consistency.
Table 1.
| Area | Pond | Latitude | Longitude | Maximum depth (cm) | pH | Temperature (°C) | Conductivity (μS/cm) | Whole water total mercury (ng/g) | Whole water methylmercury (ng/g) | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Amey | MinTill | 52° | 30′ | 14.04″ | 107° | 18′ | 04.32″ | 90 | 7.12 | 19.38 | 978 | 8.18 | 1.445 |
| Org | 52° | 29′ | 15.00″ | 107° | 21′ | 33.00″ | 50 | 6.01 | 18.38 | 202 | 4.63 | 0.214 | |
| Conv | 52° | 31′ | 03.00″ | 107° | 18′ | 23.00″ | 70 | 7.14 | 18.96 | 1026 | 12.06 | 5.396 | |
| Baumel | Wild | 52° | 10′ | 45.80″ | 105° | 15′ | 42.20″ | 230 | 8.57 | 22.87 | 789 | 3.28 | 0.840 |
| Bull | MinTill | 52° | 44′ | 19.68″ | 105° | 01′ | 17.04″ | 95 | 8.19 | 27.22 | 162 | 5.45 | 0.091 |
| Demong | Org | 52° | 33′ | 27.70″ | 105° | 35′ | 24.80″ | 200 | 7.49 | 21.08 | 386749 | 3.63 | 0.624 |
| Conv | 52° | 33′ | 27.70″ | 105° | 35′ | 24.80″ | 140 | 6.97 | 21.37 | 151 | 2.33 | 0.520 | |
| Friesen | MinTill | 52° | 42′ | 57.96″ | 106° | 29′ | 08.16″ | 45 | 7.14 | 27.7 | 270 | 4.71 | 0.258 |
| Org | 52° | 40′ | 27.00″ | 106° | 38′ | 57.00″ | 70 | 6.39 | 16.16 | 318 | 7.91 | 0.088 | |
| Gillis | MinTill | 52° | 40′ | 54.12″ | 104° | 14′ | 27.96″ | 130 | 8.04 | 20.84 | 397 | 2.66 | 1.330 |
| Conv | 52° | 47′ | 39.84″ | 103° | 52′ | 48.36″ | 99 | 8.04 | 20.898 | 1026 | 9.54 | 3.391 | |
| Leight | Wild | 52° | 28′ | 11.00″ | 104° | 32′ | 11.00″ | 75 | 8.9 | 23.34 | 941 | 7.13 | 2.739 |
| MinTill | 52° | 25′ | 06.00″ | 104° | 24′ | 41.00″ | 70 | 7.38 | 19.87 | 573 | 7.87 | 3.193 | |
| Org | 52° | 18′ | 36.00″ | 104° | 20′ | 21.00″ | 100 | 8.21 | 22.94 | 427 | 4.08 | 1.112 | |
| Loiselle | MinTill | 52° | 18′ | 05.40″ | 106° | 13′ | 18.48″ | 315 | 6.68 | 16.13 | 268 | 6.83 | 2.598 |
| Conv | 52° | 15′ | 57.00″ | 106° | 04′ | 21.00″ | 70 | 7.05 | 21.7 | 330 | 2.96 | 0.386 | |
| Mumm | Conv | 53° | 16′ | 48.00″ | 106° | 10′ | 23.16″ | 100 | 7.34 | 19.1 | 944 | No data | No data |
| Naharney | Wild | 52° | 45′ | 07.00″ | 107° | 07′ | 59.88″ | 155 | 9 | 23.2 | 1633 | 3.33 | 0.561 |
| Conv | 52° | 43′ | 47.00″ | 107° | 16′ | 04.00″ | 100 | 7.7 | 18.8 | 1240 | 5.61 | 2.084 | |
| Weiland | Wild | 52° | 14′ | 39.70″ | 104° | 57′ | 58.10″ | 85 | 8.85 | 25.8 | 688 | 6.42 | 1.289 |
| MinTill | 52° | 27′ | 34.00″ | 105° | 12′ | 52.56″ | 83 | 7.36 | 21.02 | 933 | 4.47 | 1.852 | |
| Conv | 52° | 27′ | 34.00″ | 105° | 07′ | 17.00″ | 140 | 7.97 | 21.12 | 1476 | 3.42 | 1.766 | |
Note: Wild, wildlife area; MinTill, minimum tillage farms; Org, organic farms; Conv, conventional farms.
Fig. 1.
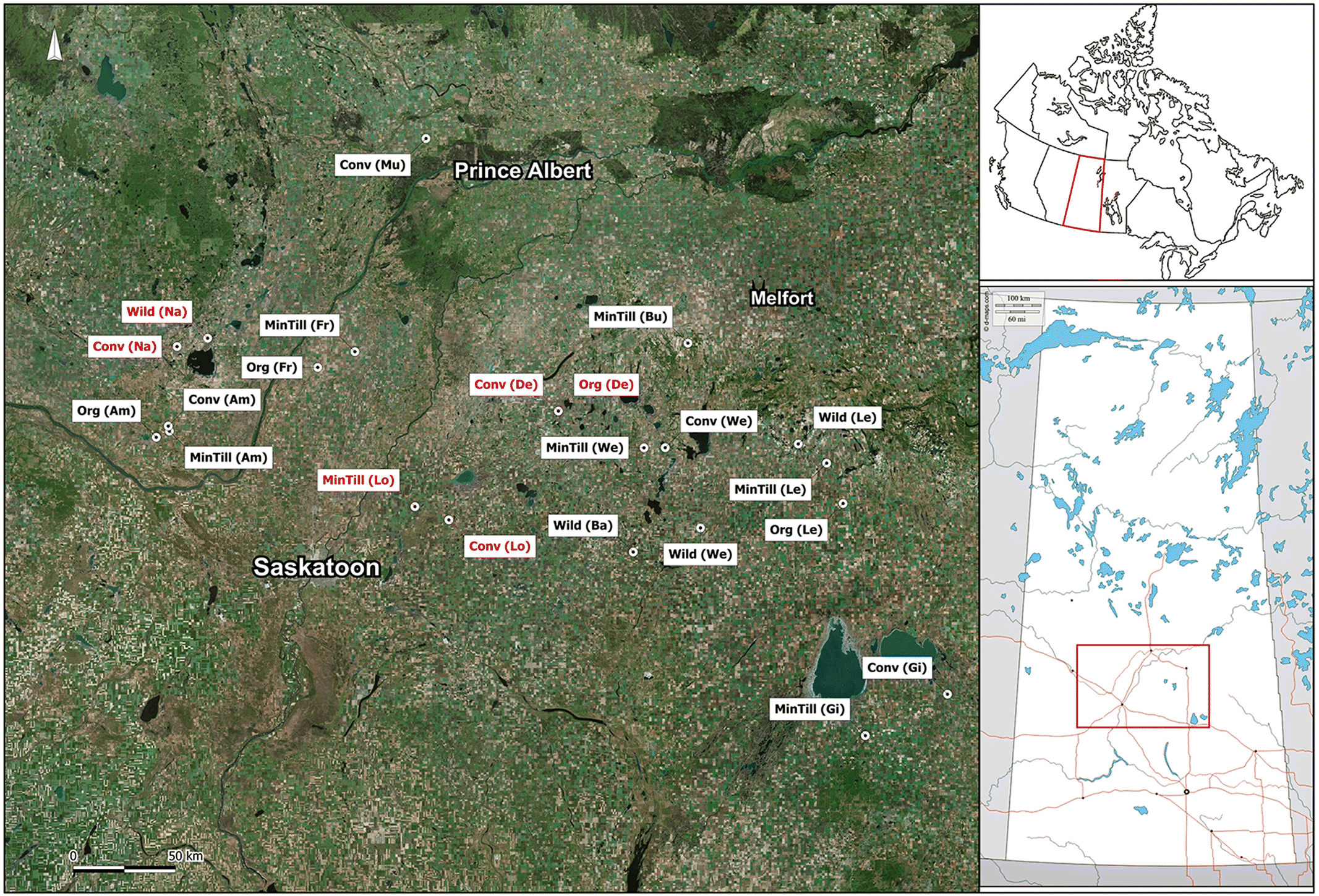
{kind=link}
Sample collection
Tadpoles
Boreal chorus frogs and wood frogs are common across the entire geographic area. Tadpoles were collected opportunistically during the day within each pond from 14 to 18 June 2010. Dip nets were used to collect tadpoles in the shallow water nearest to the shore around the perimeter of each wetland pond for 30 min. All tadpoles collected were separated into plastic containers where they were visually identified according to Preston (1982). A subsample of three boreal chorus frog and three wood frog tadpoles were randomly selected, euthanized using buffered tricaine methane/sulfonate (MS-222), rinsed in ultrapure water, placed in separate Ziploc® bags, flash frozen with dry ice, and stored in the field on dry ice. Size was not a criterion for tadpole selection. If three frogs could not be collected in 30 min, one or two frogs were taken instead. Euthanization of frogs was performed using procedures approved by the Canadian Council on Animal Care (2010) with appropriate permits. We obtained a total of 32 boreal chorus frog and 61 wood frog tadpoles from 20 different wetland sites. Developmental stage (using Gosner 1960) was recorded prior to sample preparation for THg analysis (see below). Tadpoles were stored in a laboratory freezer (−14 °C) until analysis.
Water samples
Water samples for both standard water quality parameters and THg and MeHg concentration were collected by wading into the center of the pond to chest height. A “clean-hands–dirty-hands” technique was used to collect unfiltered water samples into precleaned glass bottles for Hg analysis (St. Louis et al. 1994). Hg samples were preserved by acidification using trace metal grade HCl (at 0.2% and 0.4% by volume for THg and MeHg, respectively). Water chemistry samples were collected concurrently, filtered using 0.45 μm glass fiber filters, and stored at 4 °C until analysis. Wetland ponds were sampled for conductivity (mS/cm), pH, and water temperature (°C) using a YSI 556 multiprobe sonde. Maximum depth (cm) was estimated at the center of each wetland pond using the cord of the probe, which was marked every 25 cm.
Sample preparation and Hg analysis
We used THg as an indication of potential MeHg uptake of tadpoles in this study. A relationship between THg and MeHg concentration in both adult and larval amphibians has been previously demonstrated (%MeHg in tadpoles in other studies ranged from 7% to 61% (Bergeron et al. 2010a; Loftin et al. 2012)); however, the use of THg as a proxy for MeHg is generally not appropriate if %MeHg is less than 90%. As a result, we used THg as an indication of potential MeHg uptake, with caution, in the context of possible sources of Hg to higher trophic level organisms.
Samples were freeze-dried, and whole tadpoles (ranging from dry weights of 5.8–73.5 mg and 70–450 mg for boreal chorus frog and wood frog tadpoles, respectively) were individually digested in Teflon bombs with a metal grade nitric:sulfuric acid ratio of 7:3 at 125 °C for 2 h. Samples were then further digested in 9 mL of ultrapure water and 1 mL of bromine monochloride at 60 °C overnight. Aliquots (1–2 mL dependent on the estimated THg concentration of the sample) were then analyzed for THg using a Tekran® Series 2600 Cold Vapor Atomic Fluorescence Spectrophotometer (United States Environmental Protection Agency 2002). Recoveries of certified reference material (National Research Council DORM-3 for THg) were between 99.6% and 100% of the certified values. The concentration of THg in the water samples was similarly analyzed following EPA Method 1631 (United States Environmental Protection Agency 2002). MeHg concentration was quantified using a Tekran 2700 at the University of Western Ontario, following distillation on a Tekran 2750 at the University of Regina. Spike recoveries on acid blanks and spike samples were between 97% and 109% and 101% and 112%, respectively. Minimum detection limits were less than 0.3 ng/L for THg and 0.02 ng/L for MeHg. Approximately 10% of samples were analyzed in duplicate.
Water chemistry analysis
Water samples were analyzed for concentrations of DOC, total nitrogen, anion (Cl, F, and SO4), cation (Na, Ca, Mg, and K), total dissolved solid and total phosphorus concentrations, and turbidity using standard methods (Stainton et al. 1977). Anion and cation data are presented as total concentrations of all anions and cations.
Statistical analysis
All data are presented in both concentration (ng THg/g dry body tissue ± standard error) and body burden (ng THg/dried frog ± standard error). We analyzed both THg concentration and body burden in whole tadpoles using linear mixed models (package lme4 in R version 3.1.1; R Core Team 2016). Both response variables were ln-transformed prior to analysis as the original data spanned 2–3 orders of magnitude and showed strong heteroskedasticity. We developed a set of eight models for each response variable that included all subsets of terms for species, metamorphic stage, and body mass as fixed effects. Each model also included a random effect for pond. We ranked models using the second-order Akaike information criterion (AIC), then calculated model weights (ω i ) as a measure of evidence for each model having the best predictive ability from among those considered. We assessed the relative importance of each predictor variable based on the sum of model weights (Σω i ) for all models in which it was included: variables with Σω i approaching one were very likely to improve model fit, those with Σω i near zero were very unlikely to improve fit, and those with intermediate Σω i had mixed support (Burnham and Anderson 2002).
We performed a principal components analysis (PCA) to explore how patterns of MeHg variation among ponds were related to water chemistry. All variables except for water temperature, depth, and pH were log-transformed prior to analysis. We substituted the mean values for missing observations for three cases in which ponds did not have data for total dissolved solids, MeHg, or THg; results using these substitutions were not noticeably different from those obtained by removing ponds with missing data from the PCA.
Results and discussion
Differences in THg in wood frog vs. boreal chorus frog tadpoles
Body burden and THg concentration in all tadpoles ranged from 0.5 to 2300.5 ng/frog dry weight (dw) and 25.9 to 7889.3 ng/g dw, respectively (Tables S1 and S2). Lower values were not outside the range of those measured in other studies; however, upper values exceeded those of any previous study, including in amphibians from some contaminated studies (see Bradford et al. 2012 for a summary). The variables species, stage, and mass explained 21% of total variation for body burden in our models but had negligible predictive ability for THg concentration (Table S3). Nearly all the remaining variation in both body burden and THg concentration was attributable to differences among ponds (R 2 = 96% when including the random pond effect in both cases; Table S3).
The THg body burden of boreal chorus frog tadpoles (n = 32, mean ± standard error: 33 ± 17 ng/frog) was lower than that of wood frog tadpoles (n = 61, mean = 329 ± 76 ng; Fig. 2a). THg concentration showed a similar trend (mean = 597.2 ± 132.1 ng/g and 1668.9 ± 351.6 ng/g for boreal chorus frogs and wood frogs, respectively; Fig. 2b). Bioaccumulation of MeHg through trophic levels has been well documented, as prey items are the primary source of MeHg for aquatic organisms (Hall et al. 1997; Rimmer et al. 2005; Ward et al. 2010). Boreal chorus frog tadpoles are herbivorous and feed on algae, whereas wood frog tadpoles are omnivorous, and likely supplement a primary diet of algae with invertebrates (e.g., Odonata and Dytiscidae) and eggs of amphibians including wood frog tadpoles (McDiarmid and Altig 1999; Wells 2007). Thus, the lower values observed in boreal chorus frogs were not surprising considering the trophic position of both species.
Fig. 2.

There was also high variation within species within ponds, especially for wood frog tadpoles. This may be due to life history parameters such as stage of development, because greater development leads to more opportunity for Hg biomagnification and a change in diet as the organism approaches metamorphosis (Bradford et al. 2012). To address differences in development timelines, we tested the effect of metamorphic stage on body burden and THg concentration. Models that included both species and stage were unambiguously supported over other models for body burden (Σω i > 0.999 for both variables; Table 2), providing further evidence of Hg bioaccumulation with metamorphic stage. There was also some evidence that tadpoles with greater mass had a higher body burden, after accounting for effects of both species and metamorphic stage (Σω i > 0.77; Table 2). We could not definitively conclude that any of the predictors had an effect on THg concentration (Σω i = 0.661, 0.727, and 0.407 for species, stages, and mass, respectively), but there was some evidence that more developed wood frogs had a higher THg concentration than less developed boreal chorus frogs (Fig. S1).
Table 2.
| Response | Predictors | ΔAICC | ω i |
|---|---|---|---|
| Natural logarithm of total mercury concentration (ng/g) | |||
| Species, stage | 0.00 | 0.40 | |
| Stage, mass | 0.95 | 0.25 | |
| Species | 1.66 | 0.17 | |
| Mass | 3.45 | 0.07 | |
| Species, stage, mass | 3.74 | 0.06 | |
| Species, mass | 5.39 | 0.03 | |
| Stage | 6.07 | 0.02 | |
| Natural logarithm of body burden (ng/frog) | |||
| Species, stage, mass | 0.00 | 0.77 | |
| Species, stage | 2.43 | 0.23 | |
Note: ω i , model weight.
Given the lack of study of Hg in amphibians in prairie wetlands, we decided to compare our results with lower trophic organisms from other studies in similar systems. Gastropods and some Corixidae feed on algae and are therefore at a similar trophic level as boreal chorus frog tadpoles (Clarke 1981; Merritt and Cummins 1996). The THg concentration in pond snails (Gastropods and Lymnaeidae) and aquatic insects (Corixidae, Odonata, Dytiscidae, and Notonectidae) from wetlands in the PPR ranged from 11.7 to 161.2 ng/g (Bates 2010) and 9.7 and 507.1 ng/g (Bates and Hall 2012), respectively. These values were lower than our boreal chorus frog tadpoles (25.9–2736.4 ng/g). The potentially higher accumulation in tadpoles compared with other organisms at similar trophic levels suggests that tadpoles may be an important vector for Hg within aquatic and terrestrial food webs as the Hg accumulated by tadpoles is retained throughout metamorphosis (Bergeron et al. 2010a). Elevated THg levels may be problematic in areas where tadpoles are abundant prey sources for higher trophic levels, as tadpoles may increase the amount of MeHg accumulated through food webs.
Differences in THg in tadpoles in different ponds
After accounting for species, stage, and weight, we observed significant pond-to-pond variability in both body burden and concentration (Table S3). Tadpoles from wetland ponds in three areas (labelled as Demong, Loiselle, and Naharney) had THg body burdens and concentrations that were an average of 60 and 38 times higher, respectively, than tadpoles from ponds in other areas (Figs. 3a, 3b). These very large differences dwarfed variation in THg among the remaining ponds.
Fig. 3.

Why were concentrations in these ponds so high?
Factors that influence HgII methylation and MeHg bioavailability are important controls in MeHg bioaccumulation in aquatic organisms (Wiener 2010; Li and Cai 2013; Hsu-Kim et al. 2013; Paranjape and Hall 2017). We explored water chemistry and physical parameters that are often useful in explaining the variation of MeHg and THg in whole water (such as DOC, pH, total anion concentrations; see Methods section for complete list), and thus Hg in aquatic animals, using PCA (Fig. 4). We did not identify any water chemistry parameters, including THg and MeHg concentrations in water, as being strongly associated with the concentration of Hg in tadpoles in our systems.
Fig. 4.

Differences in Hg concentration in the ponds were apparent for both species of tadpoles, suggesting that Hg concentration in algae and lower trophic organisms must also have been elevated because diet is the main vector of Hg in most vertebrates (Burger et al. 2014). Another explanation of high values in certain ponds is that the gastrointestinal tracts (GT) of tadpoles sampled from the high Hg ponds contained sediment with an elevated Hg concentration. Burger and Snodgrass (1998) showed that the concentration of Hg in tadpoles GTs was elevated compared with Hg concentration in the tail or rest of the body, suggesting that gut contents can contribute to overall body concentration. In addition, tadpoles have been shown to accumulate metals from sediment only (Snodgrass et al. 2004). We did not measure THg concentration in sediments in our ponds. However, our previous work has shown that Hg concentration in sediments can vary in these types of systems (Hoggarth et al. 2015), and it is therefore conceivable that the high Hg ponds had elevated Hg concentration in the sediments. If sediments were a significant source of Hg for tadpoles, we would expect to find a high inorganic Hg to MeHg ratio in the organisms. A further exploration of Hg speciation in tissues is required, and clearly a more thorough study on the biogeochemical cycling of Hg, including concentration in the sediments, in these high Hg ponds is warranted.
Wetland geomorphology is likely important in the determination of Hg levels in aquatic organisms. For example, natural areas tend to retain more water in soils and thus input less water into wetlands compared with agricultural areas (van der Kamp et al. 2003), leading to a greater diversity in hydrological patterns, especially in prairie wetland ponds (Hayashi et al. 2016). Wet–dry cycling is important for the methylation process as it reoxidizes sulfur for metabolism by sulfate reducing bacteria, and therefore, methylation of HgII, and subsequent bioaccumulation of MeHg in biota, may be influenced by the frequency of drying and flooding in wetlands (Sando et al. 2007; Eckley et al. 2015; Singer et al. 2016). Consequently, intermittent wetlands will contain higher amounts of MeHg in the water and in residing fish (Ackerman and Eagles-Smith 2010; Coleman Wasik et al. 2015). Second, the vegetation composition of riparian zones may also be an important factor in determining MeHg levels in aquatic habitats because a higher amount of shading by riparian zones is associated with higher MeHg levels in aquatic organisms (Balogh et al. 2002; Ward et al. 2010). Riparian vegetation of our ponds was typical of PPR wetlands and could provide shading of sediments where methylation could be occurring. These two examples demonstrate that the processes controlling Hg accumulation in these tadpoles are much more complex than inorganic Hg deposition alone.
Conclusions
Hg cycling in the wetland ponds of the PPR is vastly understudied. We present the first data set on Hg in frog tadpoles from this region. Because wood frog tadpoles had a higher THg concentration and body burden than boreal chorus frog tadpoles, our results supported our hypothesis that biomagnification of Hg occurred with increasing trophic level in tadpoles in the wetland ponds of the PPR. Both boreal chorus frog and wood frog tadpoles contained relatively high THg levels compared with invertebrates at similar trophic levels in the PPR and tadpoles in other studies. After accounting for species, stage, and weight, we observed high pond-to-pond variability in both body burden and concentration (Table S3). Tadpoles from ponds in three areas had an ∼40× higher THg body burden and concentration than those from other ponds, and these differences were not related to any water chemistry or physical parameter measured. Differences in wetland geomorphology likely played an important role in the determination of Hg in aquatic organisms. Elevated Hg concentration is potentially problematic for both the tadpole populations and tadpole predators due to the toxicological impacts of Hg bioaccumulation, and tadpoles may be important vectors of Hg from aquatic to terrestrial systems.
Acknowledgements
This work was supported by a National Science and Engineering Research Council Discovery Grant to BDH. Environment Canada provided water chemistry data. We thank J Kindrachuk, C Hoggarth, J Poissant, Dr. B Parker, and Environment Canada sampling crew for assistance in sampling, analysis, and initial editing of the manuscript. We are especially indebted to Dr. D Donald who shared the experimental design, site locations, sampling help, and expertise. Thank you to the many landowners who provided access to their land. Appropriate permits and protocols for animal care were obtained from the University of Regina Animal Utilization Protocol 11-04.
References
Ackerman JT, and Eagles-Smith CA. 2010. Agricultural wetlands as a potential hotspot for mercury bioaccumulation: experimental evidence using caged fish. Environmental Science and Technology, 44: 1451–1457.
Balogh SJ, Huang Y, Offerman HJ, Meyer ML, and Johnson DK. 2002. Episodes of elevated methylmercury concentrations in prairie streams. Environmental Science and Technology, 36: 1665–1670.
Bank MS, Burgess JR, Evers DC, and Loftin CS. 2007a. Mercury contamination of biota from Acadia National Park, Maine: a review. Environmental Monitoring and Assessment, 126: 105–115.
Bank MS, Croker J, Connory B, and Amirbahman A. 2007b. Mercury bioaccumulation in green frog (Ranas clamitans) and bullfrog (Rana catesbeiana) tadpoles from Acadia National Park, Maine, USA. Environmental Toxicology and Chemistry, 26: 118–125.
Bates LM. 2010. Investigating the bioaccumulation of methylmercury in invertebrates from Saskatchewan Prairie Pothole wetlands. M.Sc. thesis, University of Regina, Regina, Saskatchewan. 101 p.
Bates LM, and Hall BD. 2012. Concentrations of methylmercury in invertebrates from wetlands of the Prairie Pothole Region of North America. Environmental Pollution, 160: 153–160.
Batt BD, Anderson MG, Anderson CD, and Caswell FD. 1989. The use of prairie potholes by North American ducks. In Northern prairie wetlands. Edited by AG Van der Valk. Iowa State University Press, Ames, Iowa. pp. 204–227.
Batzer DP, and Sharitz RR. 2006. Ecology of freshwater and estuarine wetlands. University of California Press, Berkley, California.
Bergeron CM, Bodinof CM, Unrine JM, and Hopkins WA. 2010a. Bioaccumulation and maternal transfer of mercury and selenium in amphibians. Environmental Toxicology and Chemistry, 29: 989–997.
Bergeron CM, Bodinof CM, Unrine JM, and Hopkins WA. 2010b. Mercury accumulation along a contamination gradient and nondestructive indices of bioaccumulation in amphibians. Environmental Toxicology and Chemistry, 29: 980–988.
Boening DW. 2000. Ecological effects, transport, and fate of mercury: a general review. Chemosphere, 40: 1335–1351.
Bradford DF, Kramer JL, Gerstenberger SL, Tallent-Halsell NG, and Nash MS. 2012. Mercury in tadpoles collected from remote alpine sites in the southern Sierra Nevada mountains, California, USA. Archives of Environmental Contamination and Toxicology, 62: 135–140.
Branfireun BA, Roulet NT, Kelly CA, and Rudd JWM. 1999. In situ sulphate stimulation of mercury methylation in a boreal peatland: toward a link between acid rain and methylmercury contamination in remote environments. Global Biogeochemical Cycles, 13: 743–750.
Burger J, and Snodgrass J. 1998. Heavy metals in bullfrog (Rana catesbeiana) tadpoles: effects of depuration before analysis. Environmental Toxicology and Chemistry, 17, 2203–2209.
Burger J, Feinberg J, Jeitner C, Gochfeld M, Donio M, and Pittfield T. 2014. Selenium:mercury molar ratios in bullfrog and leopard frog tadpoles from the northeastern United States. Ecohealth, 11, 154–163.
Burnham KP, and Anderson DR. 2002. Model selection and multimodel inference: a practical information-theoretic approach. Springer, New York, New York.
Canadian Council on Animal Care. 2010. Guide to the care and use of experimental animals [online]: Available from ccac.ca/en_/standards/guidelines.
Clarke AH. 1981. The freshwater molluscs of Canada. National Museum of Natural Sciences, National Museums of Canada, Ottawa, Ontario. 446 p.
Coleman Wasik JK, Engstrom DR, Mitchell CPJ, Swain EB, Monson BA, Balogh SJ, et al. 2015. The effects of hydrologic fluctuation and sulfate regeneration on mercury cycling in an experimental peatland. Journal of Geochemical Research: Biogeosciences, 120: 1697–1715.
Corn PS. 2000. Amphibian declines: review of some current hypotheses. In Ecotoxicology of amphibians and reptiles. Edited by DW Sparling, G Linder, and CA Bishop. SETAC Press, Pensacola, Florida. pp. 663–696.
Donald DB, Gurprasad NP, Quinnett-Abbott L, and Cash K. 2001. Diffuse geographic distribution of herbicides in northern prairie wetlands. Environmental Toxicology and Chemistry, 20: 273–279.
Eckley CS, Luxton TP, McKernan JL, Goetz J, and Goulet J. 2015. Influence of reservoir water level fluctuations on sediment methylmercury concentrations downstream of the historical Black Butte mercury mine, OR. Applied Geochemistry, 61: 284–293.
Environment Canada. 2016. Canadian Climate Normals 1981–2010 [online]: Available from climate.weather.gc.ca/climate_normals/results_1981_2010_e.html?searchType=stnProv&lstProvince=SK&txtCentralLatMin=0&txtCentralLatSec=0&txtCentralLongMin=0&txtCentralLongSec=0&stnID=3002&dispBack=0.
Ficken KLG, and Byrne PG. 2013. Heavy metal pollution negatively correlates with anuran species richness and distribution in south-eastern Australia. Austral Ecology, 38: 523–533.
Gilmour CC, and Henry EA. 1991. Mercury methylation in aquatic systems affected by acid deposition. Environmental Pollution, 71: 131–169.
Gilmour CC, Podar M, Bullock AL, Mitchell Grahan A, Brown S, Somenahally AC, et al. 2013. Mercury methylation by novel microorganisms from new environments. Environmental Science and Technology, 47: 11810–11820.
Gosner KL. 1960. A simplified table for staging anuran embryos and larvae with notes on identification. Herpetologica, 16: 183–190.
Graydon JA, Louis VLS, Hintelmann H, Lindberg SE, Sandilands KA, Rudd JWM, et al. 2008. Long-term wet and dry deposition of total and methyl mercury in the remote Boreal ecoregion of Canada. Environmental Science and Technology, 42: 8345–8351.
Hall BD, Baron LA, and Somers CM. 2009. Mercury concentrations in surface water and harvested waterfowl from the Prairie Pothole Region of Saskatchewan. Environmental Science and Technology, 43: 8759–8766.
Hall BD, Bodaly RA, Fudge RJP, Rudd JWM, and Rosenberg DM. 1997. Food as the dominant pathway of methylmercury uptake by fish. Water, Air, and Soil Pollution, 100: 13–24.
Hall BD, Manolopoulos H, Hurley JP, Schauer JJ, St. Louis VL, Kenski D, et al. 2005. Methyl and total mercury in precipitation in the Great Lakes region. Atmospheric Environment, 39: 7557–7569.
Hayashi M, Van der Kamp G, and Rosenberry DO. 2016. Hydrology of prairie wetlands: Understanding the integrated surface-water and groundwater processes. Wetlands, 36(suppl 2): 237–254.
Hightower JM. 2009. Diagnosis: mercury. Island Press, Washington, DC. 328 p.
Hintelmann H. 2010. Organomercurials: their formation and pathways in the environment. Metal Ions in Life Sciences, 7: 365–401.
Hoggarth CGJ, Mitchell CPJ, and Hall BD. 2015. Mercury methylation in high and low-sulphate impacted wetland ponds within the Prairie Pothole Region of North America. Environmental Pollution, 205: 269–277.
Hothem RL, Jennings MR, and Crayon JJ. 2010. Mercury contamination in three species of anuran amphibians from the Cache Creek watershed, California, USA. Environmental Monitoring Assessment, 163: 433–448.
Hsu-Kim H, Kucharzyk KH, Zhang T, and Deshusses MA. 2013. Mechanisms regulating mercury bioavailability for methylating microorganisms in the aquatic environment: a critical review. Environmental Science and Technology, 47: 2441–2456.
Johnson WC, Werner B, Guntenspergen GR, Voldseth RA, Millett BV, Naugle DE, et al. 2010. Prairie wetland complexes as landscape functional units in a changing climate. BioScience, 60: 128–140.
Li Y, and Cai Y. 2013. Progress in the study of mercury methylation and demethylation in aquatic environments. Chinese Science Bulletin, 58: 177–185.
Loftin CS, Calhoun AJK, Nelson SJ, Elskus AA, and Simon K. 2012. Mercury bioaccumulation in wood frogs developing in seasonal pools. Northeastern Naturalist, 19: 579–600.
McDiarmid RW, and Altig R. 1999. Tadpoles: the biology of anuran larvae. University of Chicago Press, Chicago, Illinois.
Merritt RW, and Cummins KW. 1996. An introduction to the aquatic insects of North America. Kendall-Hunt Publishing Company, Dubuque, Iowa.
Morel FMM, Kraepiel AML, and Amyot M. 1998. The chemical cycle and bioaccumulation of mercury. Annual Review of Ecology and Systematics, 29: 543–566.
Paranjape AR, and Hall BD. 2017. Recent advances in the study of mercury methylation in aquatic systems. FACETS, 2: 85–119.
Parks JM, Johs A, Podar M, Bridou R, Hurt JRA, Smith S, et al. 2013. The genetic basis for bacterial mercury methylation. Science, 339: 1332–1335.
Preston W. 1982. The amphibians and reptiles of Manitoba. Manitoba Museum of Man and Nature, Winnipeg, Manitoba.
R Core Team. 2016. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria [online]: Available from R-project.org/.
Rimmer CC, McFarland KP, Evers DC, Miller EK, Aubry Y, Busby D, et al. 2005. Mercury concentrations in Bicknell’s thrush and other insectivorous passerines in montane forests of northeastern North America. Ecotoxicology, 14: 223–240.
Sando SK, Krabbenhoft DP, Johnson KM, Lundgrean RF, and Emerson DG. 2007. Mercury and methylmercury in water and bottom sediments of wetlands at Lostwood National Wildlife Refuge, North Dakota, 2003–2004. USGS Scientific Investigations Report 2007–5219, USGS Scientific Investigations Report 2007–5219, 74.
Scheuhammer AM, Meyer MW, Sandheinrich MB, and Murray MW. 2007. Effects of environmental methylmercury on the health of wild birds, mammals, and fish. Ambio, 36: 12–19.
Schuster PF, Krabbenhoft DP, Naftz DL, Cecil LD, Olson ML, DeWild JF, et al. 2002. Atmospheric mercury deposition during the last 270 years: a glacial ice core record of natural and anthropogenic sources. Environmental Science and Technology, 36: 2303–2310.
Singer MB, Harrison LR, Donovan PM, Blum JD, and Marvin-DiPasquale M. 2016. Hydrologic indicators of hot spots and hot moments of mercury methylation potential along river corridors. Science of the Total Environment, 568: 697–711.
Snodgrass JW, Hopkins WA, Broughton J, Gwinn D, Baionno JA, and Burger J. 2004. Species-specific responses of developing anuras to coal combustion wastes. Aquatic Toxicology, 66: 171–182.
St. Louis VL, Rudd JWM, Kelly CA, Beaty KG, Bloom NS, and Flett RJ. 1994. Importance of wetlands as sources of methyl mercury to boreal forest ecosystems. Canadian Journal of Fisheries and Aquatic Sciences, 51: 1065–1076.
Stainton MP, Capel MJ, and Armstrong FAJ. 1977. The chemical analysis of fresh water. Fisheries and Marine Services Miscellaneous Special Publications, 25: 166.
Stuart SN, Chanson JS, Cox NA, Young BE, Rodrigues ASL, Fischman DL, and Waller RW. 2004. Status and trends of amphibian declines and extinctions worldwide. Science, 306: 1783–1786.
Ugarte CA, Rice KG, and Donnelly MA. 2005. Variation of total mercury concentrations in pig frogs (Rana grylio) across the Florida Everglades, USA. Science of the Total Environment, 345: 51–59.
United States Environmental Protection Agency. 2002. Method 1631: mercury in water by oxidation, purge and trap, and cold vapor atomic fluorescence spectrometry. Revision E. EPA 821-R-95-027.
Unrine JM, Jagoe CH, Hopkins WA, and Brant HA. 2004. Adverse effects of ecologically relevant dietary mercury exposure in southern leopard frog (Rana sphenocephala) larvae. Environmental Toxicology and Chemistry, 23: 2964–2970.
Unrine JM, Hopkins WA, Romanek CS, and Jackson BP. 2007. Bioaccumulation of trace elements in omnivorous amphibian larvae: implications for amphibian health and contaminant transport. Environmental Pollution, 149: 182–192.
van der Kamp G, Hayashi M, and Gallén D. 2003. Comparing the hydrology of grassed and cultivated catchments in the semi-arid Canadian prairies. Hydrological Processes, 17: 559–575.
Ward DM, Nislow KH, and Folt CL. 2010. Bioaccumulation syndrome: identifying factors that make some stream food webs prone to elevated mercury bioaccumulation. Annals of the New York Academy of Sciences, 1195: 62–83.
Weiss-Penzias PS, Gay DA, Brigham ME, Parsons MT, Gustin MS, and ter Schure A. 2016. Trends in mercury wet deposition and mercury air concentrations across the US and Canada. Science of the Total Environment, 568: 546–556.
Wells KD. 2007. The ecology and behavior of amphibians. University of Chicago Press, Chicago, Illinois. 1148 p.
Wiener JG. 2010. Mercury exposed: advances in environmental analysis and ecotoxicology of a highly toxic metal. Environmental Toxicology and Chemistry, 32: 2175–2178.
Zillioux EJ, Porcella DB, and Benoit JM. 1993. Mercury cycling and effects in freshwater wetland ecosystems. Environmental Toxicology and Chemistry, 12: 2245–2264.
Supplementary material
Supplementary Material 1 (DOCX / 44.1 KB):
- Download
- 44.20 KB
Information & Authors
Information
Published In

FACETS
Volume 2 • Number 1 • January 2017
Pages: 315 - 329
Editor: Peter G. Kevan
History
Received: 3 August 2016
Accepted: 4 February 2017
Version of record online: 28 March 2017
Copyright
© 2017 Boczulak et al. This work is licensed under a Creative Commons Attribution 4.0 International License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Data Availability Statement
All relevant data are within the paper and in the Supplementary Material.
Key Words
Sections
Subjects
Authors
Author Contributions
SAB and BDH conceived and designed the study.
All performed the experiments/collected the data.
All analyzed and interpreted the data.
BDH contributed resources.
All drafted or revised the manuscript.
Competing Interests
The authors have declared that no competing interests exist.
Metrics & Citations
Metrics
Other Metrics
Citations
Cite As
Stacy A. Boczulak, Mark C. Vanderwel, and Britt D. Hall. 2017. Survey of mercury in boreal chorus frog (Pseudacris maculata) and wood frog (Rana sylvatica) tadpoles from wetland ponds in the Prairie Pothole Region of Canada. FACETS.
2: 315-329.
https://doi.org/10.1139/facets-2016-0041
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
Cited by
1. Methylmercury in subarctic amphibians: environmental gradients, bioaccumulation, and estimated flux
2. Complex Life Histories Alter Patterns of Mercury Exposure and Accumulation in a Pond-Breeding Amphibian
3. Site- and Individual-Level Contaminations Affect Infection Prevalence of an Emerging Infectious Disease of Amphibians
4. Metal accumulation varies with life history, size, and development of larval amphibians
5. Assessment of Sublethal Effects of Neonicotinoid Insecticides on the Life-History Traits of 2 Frog Species
6. Associations between environmental pollutants and larval amphibians in wetlands contaminated by energy-related brines are potentially mediated by feeding traits
7. Mercury exposure to red-winged blackbirds (Agelaius phoeniceus) and dragonfly (Odonata: Aeshnidae) nymphs in Prairie Pothole wetlands
8. Evaluating Wildlife Vulnerability to Mercury Pollution From Artisanal and Small-Scale Gold Mining in Madre de Dios, Peru