American lobster (Homarus americanus) hepatopancreas transcriptome reveals the significance of chitin-related genes during impoundment shell disease
Abstract
American lobsters (Homarus americanus) stored in open tidal pounds can develop impoundment shell disease (ISD), resulting in decreased marketability of the lobsters on the live market. Little is known about ISD or the immunological responses of lobsters exhibiting this disease. The objective of this project was to identify genes from H. americanus hepatopancreas that are differentially expressed in response to ISD. Lobsters were separated into asymptomatic, moderately symptomatic, and severely symptomatic groups, which represent animals with 0%, 5%–20%, and >20% lesion coverage of the carapace, respectively. RNA-seq analysis found that 134 genes were differentially expressed between groups (false discovery rate (FDR) < 0.05). Most, 80, of these genes were found exclusively in the comparison between moderately symptomatic and asymptomatic groups. All animals clustered in their proposed groups based on the expression of the differently expressed genes (DEGs), and the asymptomatic group clustered as an out-group. The expression of most DEGs was higher in the asymptomatic group than the others, which could be related to a stronger response against the disease or differences in individual resistance against ISD development. Among these genes, we highlight eight chitin-related genes, one α-2-macroglobulin-like gene, one acute phase serum amyloid A gene, one pseudohaemocyanin gene, and one trypsin-1-like gene.
Introduction
The American lobster (Homarus americanus) fishery is Canada’s most valuable fishery, generating $2 billion in commercial landings in 2021 in Atlantic Canada (Department of Fisheries and Oceans 2022). This fishery generates thousands of jobs in rural areas and is significant for the economy and culture of Atlantic Canada (FAO 2020). The present and future of the global crustacean fishery are influenced by several factors; however, disease is one of the greatest impediments to their long-term sustainability and growth. Shell diseases are the most common diseases that affect multiple crustacean genera globally. These diseases have been noted in crustaceans for at least 100 million years (Klompmaker et al. 2016) and affect many, if not most, crustacean species. They present as lesions of the shell but could be caused by unrelated infections. These diseases can manifest themselves in wild populations, or in captivity, and can cause the death of these animals by secondary infections (Cawthorn 2011).
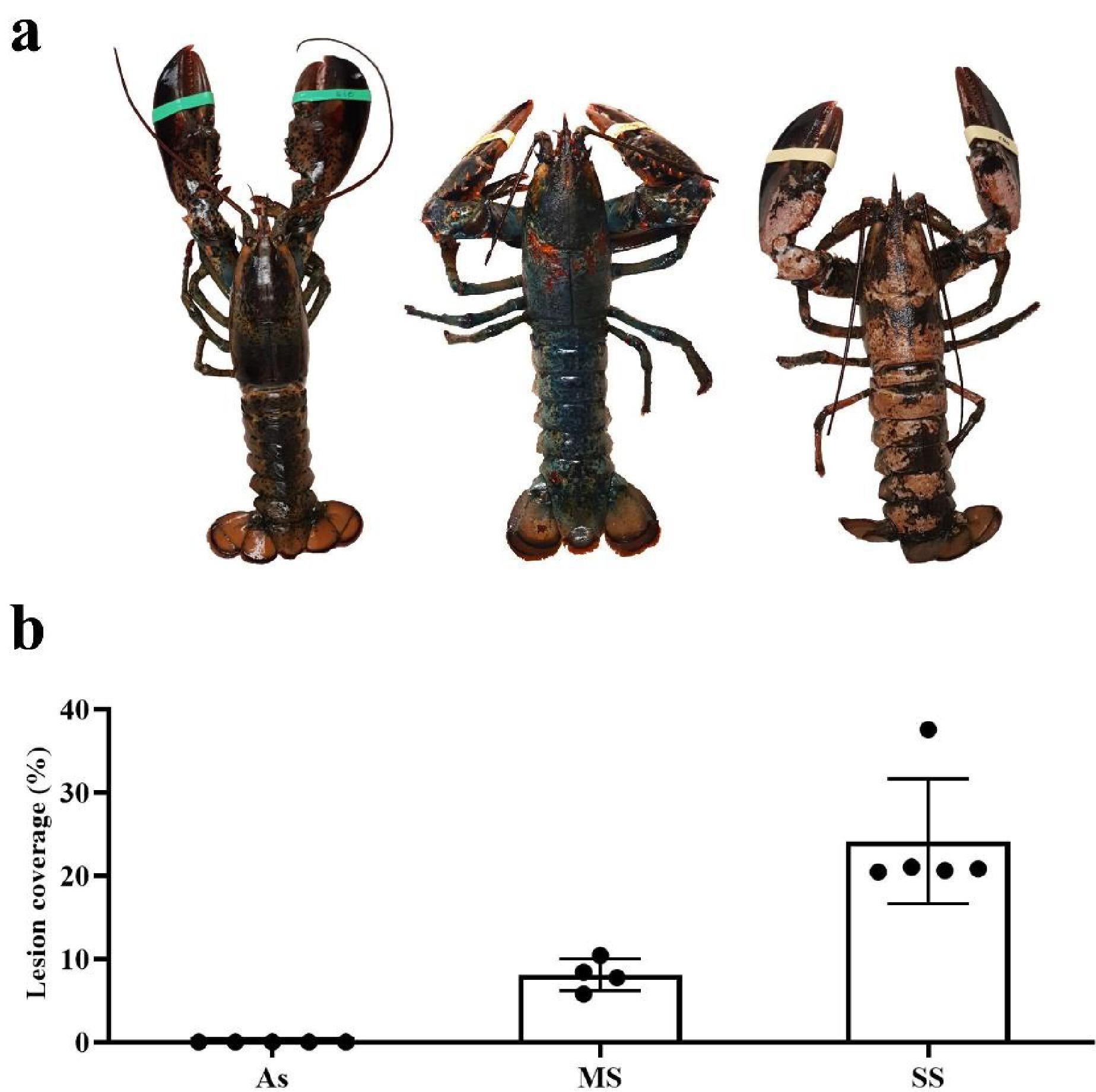
Among H. americanus, one of the most economically significant shell diseases is impoundment shell disease (ISD). ISD may occur in lobsters kept in live storage, especially during the winter (Smolowitz et al. 1992). The presence of ISD prevents animals from being sold through the live-market due to their unappealing appearance and, in more serious cases, can result in the mortality of the animal. It is characterized by the development of bilateral and symmetrical circular lesions around setal pores in the early stages of the disease, but lesions overlap and collapse to develop into a severe shell disease in advanced stages (Fig. 1a) (Theriault et al. 2008a). Although ISD was discovered in 1937, its causes were not fully characterized because many multifactorial physiological and environmental effects make identifying a single cause of shell diseases development difficult. Firstly, its development may not be due to a single microorganism, but rather a mix of effects caused by many microorganisms. Among the microorganisms found in ISD are mainly filamentous algae in early epicuticular erosions and, in advanced stages, a mix of fungi, bacteria, and protozoa (Smolowitz et al. 1992). As with other shell diseases, chitinolytic bacteria are found at all stages of the disease (Smolowitz et al. 1992). In addition to the cause of the disease not being a specific microorganism, this disease can be caused by opportunistic microorganisms from the natural microbiota of the shell when animals become more susceptible. This happens in the epizootic shell disease (ESD), a disease symptomatically similar to ISD that occurs in wild populations (Meres et al. 2012).
Fig. 1.
Like all other crustaceans, American lobsters have an innate immune system to avoid pathogenic proliferation and infections (Li and Wang 2021). The hepatopancreas is the most important immune organ in crustaceans, acting in crucial immune responses, such as phagocytosis by fixed phagocytes, and the production and release of systemic immune effectors (Ridgway et al. 2006; Sun et al. 2008; Vogt 2019; Yu et al. 2022). Little is known about the hepatopancreas immune responses during shell diseases in lobsters, where only one study evaluated the expression of hepatopancreas genes in lobsters with ESD (Tarrant et al. 2010). The advancement of high-throughput transcriptomic studies in recent years has allowed an understanding of host–pathogen interactions in more detail, which is essential to understand immune responses against complex diseases such as ISD.
The aim of this study was to identify ISD-related immune molecules by comparing the hepatopancreatic transcriptome of asymptomatic (As), moderately symptomatic (MS), and severely symptomatic (SS) lobsters. Although these lobsters were subjected to the same impoundment conditions, they developed different degrees of disease or remained As, which is possibly related to their transcriptional profiles.
Materials and methods
Homarus americanus handling and tissue collection
All research involving animals was approved, prior the beginning of the research, by the Dalhousie University Animal Care and Use Committee (approval number 1036552; January 2020) as per the Canadian Council of Animal Care guidelines. Fifteen juvenile intermolt stage American lobsters with and without visible ISD lesions were collected in March 2020 at a tidal pound operation in southwestern Nova Scotia, Canada. The lobsters were kept in aquariums containing 30 ppt seawater with constant aeration. After 48 h of acclimatation, animals were euthanized by severing the ventral nerve cord anterior to the chelae. Hepatopancreas tissue from a distal posterior lobe was macerated in TRI reagent (1.4 mol/L guanidine thiocyanate, 38% phenol, 5% glycerol, and 0.1 mol/L sodium acetate) and stored at −20 °C. Another part of the sample was fixed in Davidson’s solution (30% filtered seawater, 30% ethanol, 20% formaldehyde, 10% glacial acetic acid, and 10% glycerol) for 24 h. The fixative solution was changed to 70% ethanol after 24 h. Fixed tissues were paraffin-embedded, sliced at 5 µm, stained with haematoxylin and eosin, and examined with a light microscope to check the presence of pathogens and glycogen storage.
Disease classification
The lobsters were separated into three groups according to the percentage of ISD lesion coverage on the carapace: As group (lesion coverage = 0%), MS group (lesion coverage = 5%–20%), and SS group (lesion coverage > 20%). Leaf Doctor software was used to distinguish the disease through the color difference between healthy and unhealthy tissues. The detection threshold was adjusted to select only symptomatic tissues (Pethybridge and Nelson 2015). The percentages of lesion coverage on the cephalothorax and abdomen of the animals were used to separate the groups.
RNA extraction and sequencing
Samples were homogenized in TRI reagent and chloroform was added to the samples (1:5; v:v), and centrifuged at 12 000 × g for 15 min at 4 °C. The top aqueous phase was transferred to a new tube containing 70% alcohol (1:1; v:v). This solution was mixed and the RNA extraction was completed using the RNeasy® column extraction kit (Qiagen), according to manufacturer’s recommendations with an on-column DNase I digestion (Qiagen). Total RNA samples were eluted in molecular biology grade water and their concentration and purity were measured by spectrophotometry using a Nanodrop ND-1000. Their integrity was assessed using an RNA 6000 Nano chip on the Agilent Bioanalyzer 2100.
Total RNA samples were submitted to the Genome Quebec Innovation Center for reverse transcription, library preparation, and mRNA sequencing by Illumina NovaSeq. Sample mRNA was purified using oligo(dT)-attached beads and randomly fragmented with addition of fragmentation buffer. Reverse transcription into single-stranded cDNA was performed using mRNA template and random hexamers primers. The second-strand was synthesized by adding dNTP/dUTP, DNA polymerase I, RNase H, and custom second-strand synthesis buffer (Illumina) to the reactions. After 3′ A addition, sequencing adapter ligation and UDG degradation of the dUMP-containing strand were performed. Fragments were selected by size and PCR enriched. After quality control, libraries were sequenced on an Illumina NovaSeq sequencer to produce PE150 reads. Raw reads were deposited in NCBI-SRA database (Bioproject accession number: PRJNA1084878).
Gene expression analysis
All analyses were conducted on the usegalaxy.org HTC platform unless otherwise stated. Raw reads were trimmed using Trim-Galore (Krueger 2012) and their quality was assessed using FastQC (Andrews 2010). The reads were mapped to the H. americanus genome (GenBank accession number: GCA_018991925.1) using HISAT2 (Kim et al. 2015; Afgan et al. 2018; Polinski et al. 2021). Transcript counting was performed using StringTie and StringTie merge and the read counts was performed using featureCounts (Liao et al. 2014; Pertea et al. 2015). The normalization and gene expression comparisons between the different groups of interest were determined using DESeq2 and edgeR (Robinson et al. 2009; Love et al. 2014). The difference in gene expression was considered statistically significant when FDR < 0.05 using the Benjamini and Hochberg method. The functional annotation of differentially expressed genes (DEGs) was done using Blastx, Blast2GO, KAAS, and KEGG tools (Altschul et al. 1997; Moriya et al. 2007; Götz et al. 2008; Kanehisa et al. 2017). Heatmaps of gene expression profiles were made using heatmap.2 package (Warnes et al. 2022) in R environment (R Core Team 2023) based on log2(fold-change (FC)) of DESeq2 DEGs normalized counts comparing individuals to the average of all samples. Volcano plots were made using DESeq2 values in SRplot online platform (Tang et al. 2023).
Results
Impoundment shell disease categorization and histology
Lobsters collected from a tidal pound with ISD were separated into three groups depending on the lesion percent coverage of their carapace. The Leaf Doctor application classified six animals in the As group (0% lesion coverage), four in the MS group (lesion coverage = 8.09% ± 1.91%), and five in the SS group (lesion coverage = 24.09% ± 7.52%) (Fig. 1b). The histological examination of these animals revealed that all lobsters in this study were free from pathogens in the hepatopancreas and had similar levels of glycogen reserves (Fig. S1).
RNA sequencing of hepatopancreas tissue
Hepatopancreatic mRNA sequencing using Illumina NovaSeq PE150 resulted in 234 721 632 raw reads, which were processed into 229 870 498 filtered reads (80.47 GB), with an average of 15 324 610 ± 2654 555 clean reads per sample, (Table S1). The samples had Q20 and Q30 values equal to 97.69 ± 0.24% and 93.62 ± 0.47%, respectively.
Identification of DEGs associated with ISD
The filtered reads were used in RNA-seq pipelines using DESeq2 and edgeR to identify the DEGs in all possible comparisons between the three groups. Gene expression was quantified as log2(FC). A total of 134 and 11 DEGs were identified across all comparisons using DESeq2 and edgeR, respectively. All DEGs found using edgeR were also found using DESeq2, and for this reason, all subsequent steps were performed using the results obtained with DESeq2. From the 134 DEGs, 100 are present in the comparisons MS against As, 39 in the comparison of SS against As, and 16 in the comparison of SS against MS (Fig. 2).
Fig. 2.
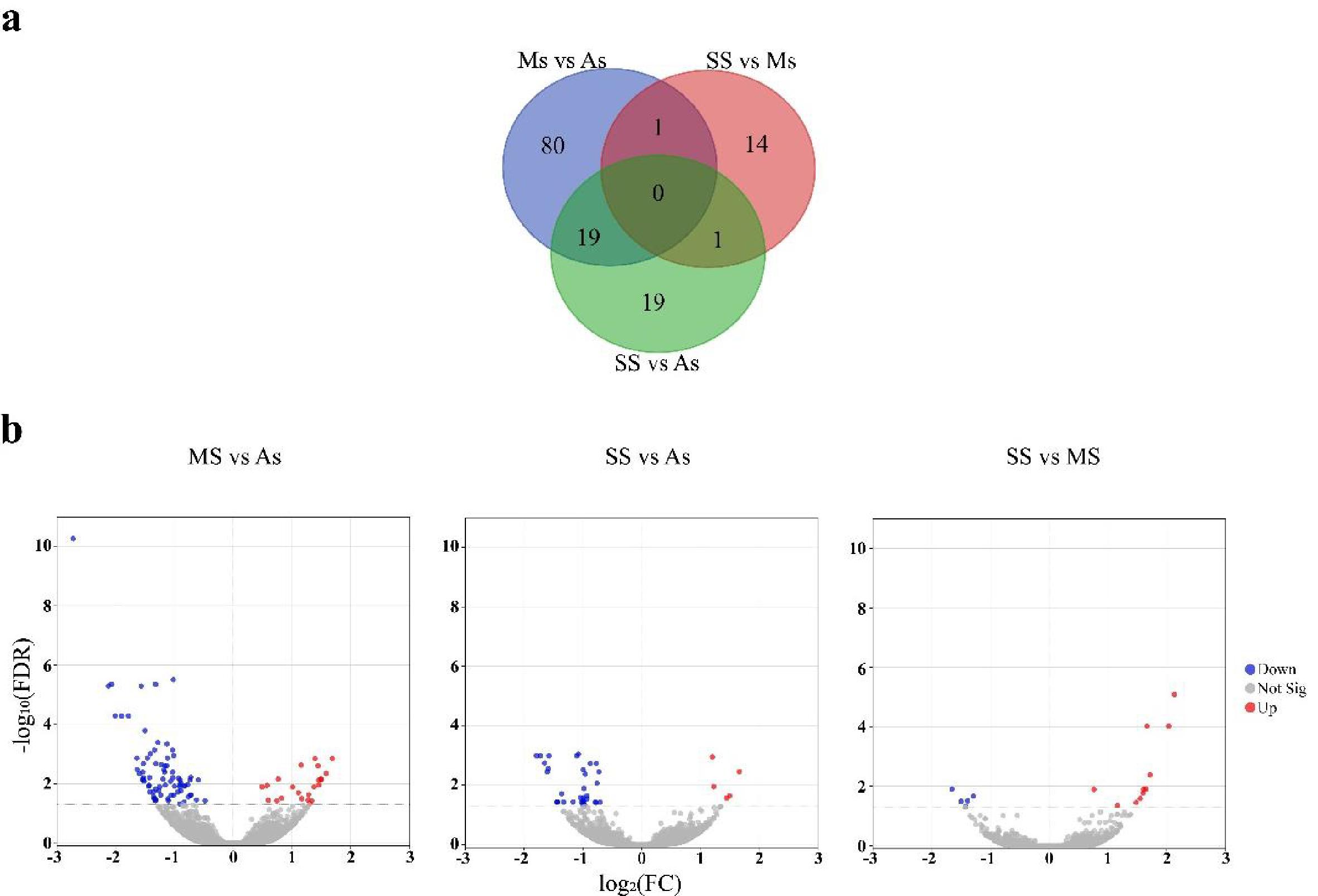
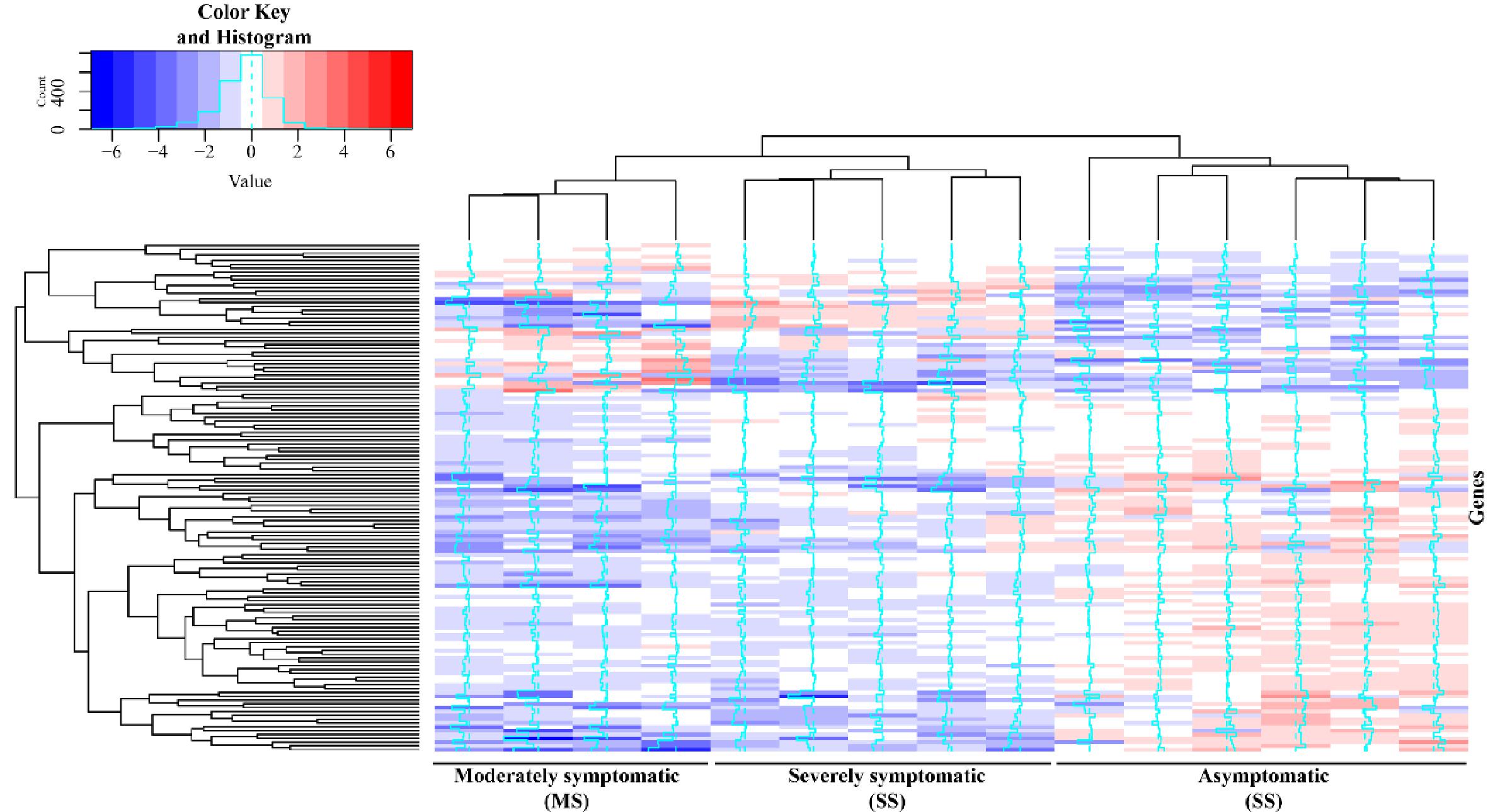
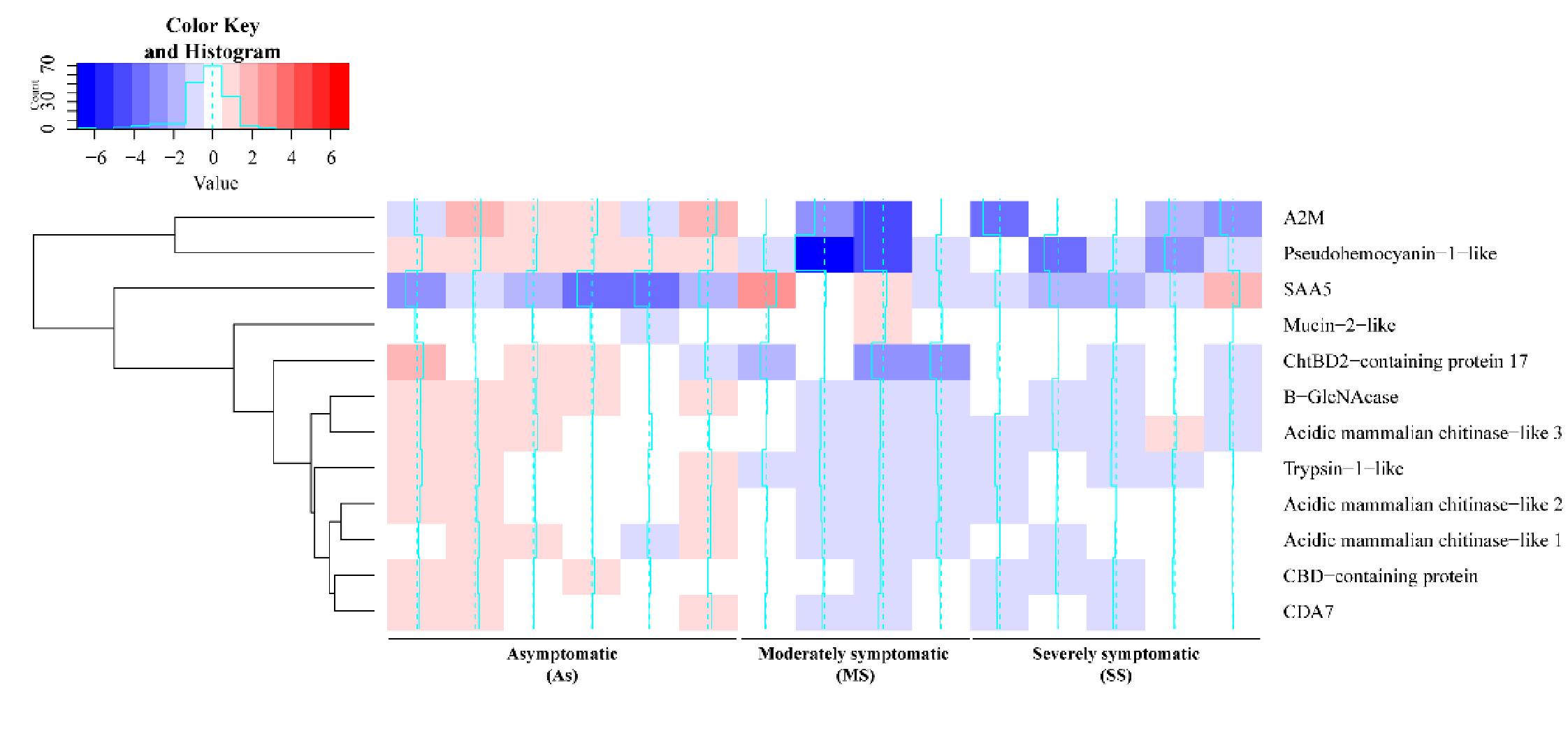
A total of 23 genes were expressed higher in MS in comparison to As, 5 higher in SS in comparison to As, and 11 higher in SS in comparison to MS (Fig. 2b). With 77, 34, and 5 genes expressed less in the same comparisons, respectively (Fig. 2b). A heatmap clustering columns (sample) and rows (genes) was made using all DEGs log2(FC) normalized counts from all samples, where log2(FC) of each gene in each sample was calculated in comparison to the average of all samples to highlight the comparative expression values (Fig. 3 and Table S2). The samples were hierarchically clustered, and the As group clustered outside the groups of symptomatic animals (Fig. 3). The transcriptional levels of most DEGs were higher in the As group compared to the others (Fig. 3).
Fig. 3.
Transcript annotation
From the 134 DEGs submitted for annotation using Blast2GO (Götz et al. 2008), 95 sequences had similarity to at least one protein in the non-redundant protein sequences database (nr) in NCBI (January 2022 database version) (Table S2). Functional annotation of the 95 sequences was performed using KAAS/KEGG, with 49 genes annotated according to their functions, but no pathways had more than three representative genes being differently expressed. Eight chitin-related DEGs were found, including three chitinases. The identified chitinases have percent identity ranging from 54.2% to 68.2% of each other, but are encoded by different genes. Their genes are positioned in different regions of the genome, present in scaffolds 16 894, 41 027, and 56 910. In fact, all chitin-related genes are in different scaffolds. In addition, other molecules potentially related to defense against infections were also found, including α-2-macroglobulin-like (A2M), pseudohaemocyanin, acute phase serum amyloid A-5 protein-like and trypsin-1-like (Figs. 4 and 5). Finally, 18 sequences had high similarity with hypothetical proteins, predicted protein, or uncharacterized protein with limited annotations.
Fig. 4.
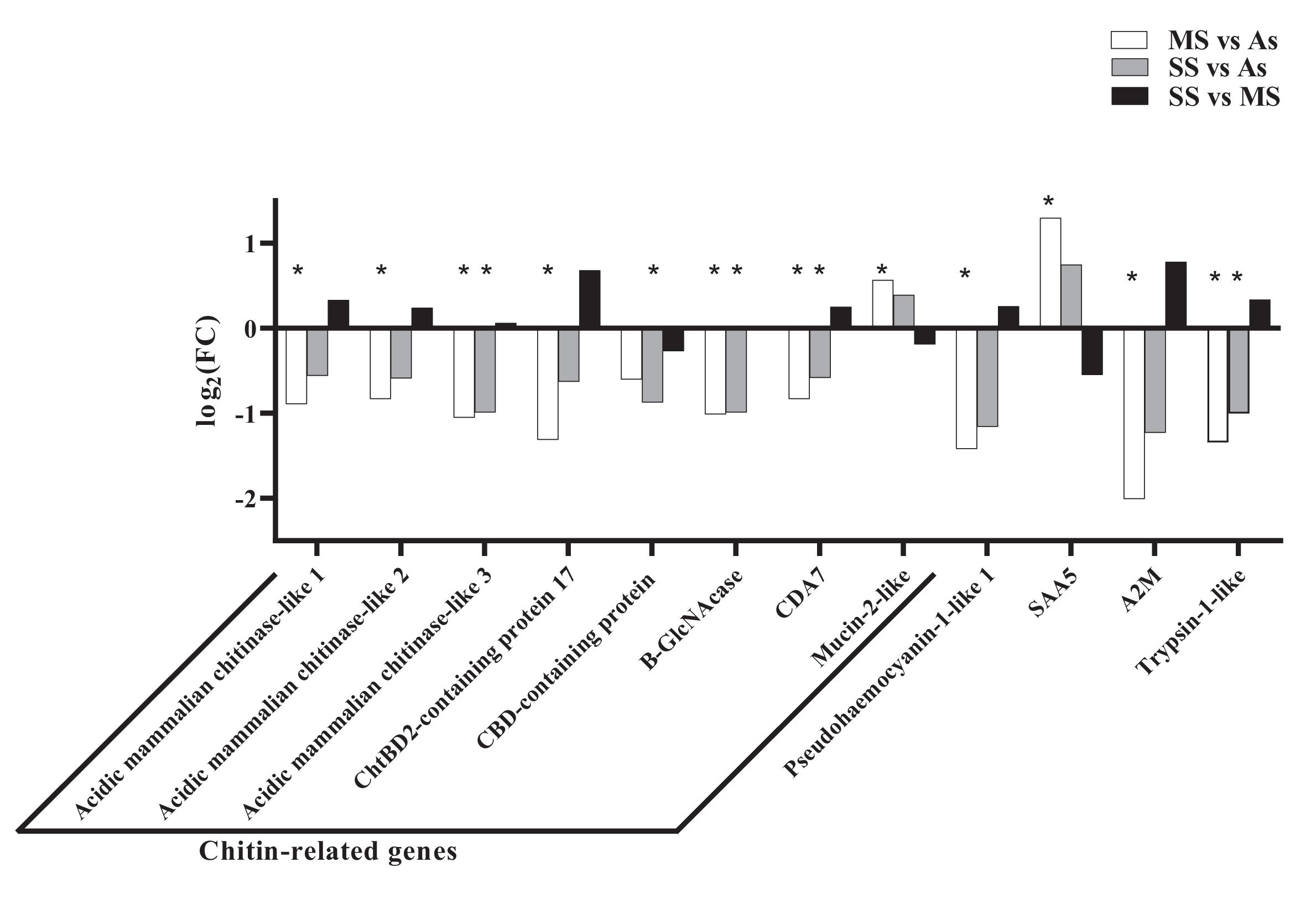
Fig. 5.
Discussion
This is the first study to use a high-throughput sequencing technique to understand H. americanus responses to ISD. The objective of this work was to characterize the genes associated with the presence of ISD in lobsters exhibiting different degrees of ISD. This work was also the first to take advantage of the first version of the recently published genome of H. americanus (Polinski et al. 2021), aligning the reads to the genome instead of a de novo transcriptome and potentially revealing genes that would not be identified in this study without access to the genome. The hepatopancreas was chosen for this study because it is the most important immune organ in crustaceans, acting in crucial immune responses (Ridgway et al. 2006; Sun et al. 2008; Vogt 2019; Yu et al. 2022). Moreover, this organ is also responsible for glycogen storage (Bonilla-Gómez et al. 2012), which is important because the animals from this study were stored for months and were unable to exhibit all of their natural behaviors. However, all animals had similar levels of glycogen reserves. Some studies showed that pathogens can be found in the haemolymph of lobsters infected with ISD (Smolowitz et al. 1992). Once in the haemolymph, these microorganisms can spread to other organs, but the hepatopancreases of the animals in the current study were free from pathogens, showing that the disease was contained at some level at this stage.
Most of the 134 DEGs potentially involved with ISD identified had higher transcriptional levels in the As group compared to the symptomatic groups (Fig. 2). After an infection, microorganisms may evade the defense mechanisms of their host in different ways, such as controlling host gene regulation (Cole and Nizet 2016). For example, Dai et al. (2022) suggest that Vibrio owensii hijacks the immune system of Fenneropenaeus merguiensis by reducing the expression of C-type lectin and ficolin, resulting in reduced recognition and phagocytosis of the microorganism. In this sense, it is possible that the largest number of genes being less expressed in symptomatic animals in the present study is due to gene suppression caused by microorganisms associated with the ISD. Alternatively, it is possible that the difference in gene expression is the result of a greater ability of As animals to have a greater constitutive immune response against ISD. All animals in this study were subjected to the same biotic and abiotic factors, where some animals developed moderately to severe degrees of the disease, others remained As. Tlusty et al. (2007) developed a hypothesis that it is the internal conditions of the lobster that make the animal resistant or susceptible to ESD, although several environmental and pathological factors also contribute to its development. Therefore, the transcriptional profiles observed in the present study could be part of the internal conditions of these animals contributing to the resistance or susceptibility to ISD development. Transcriptional profiles were previously associated with resistance and susceptibility against diseases in shrimp (de Lorgeril et al. 2005). In line with this idea, Theriault et al. (2008b) examined physiological parameters of H. americanus prior the storage in pounds and evaluated the development of ISD for 120 days. Interestingly, lobsters with lower levels of total protein developed more ISD compared to lobsters with higher levels of total protein (Theriault et al. 2008b).
Among the DEGs found in this study are several chitin-related genes, such as three chitinases and one chitooligosaccharidolytic β-N-acetylglucosaminidase-like (B-GlcNacase). While chitinases degrade the chitin molecule into chitin-oligosaccharides (GlcNAc)n, B-GlcNacase is responsible for cleaving these oligosaccharides into monomers of N-acetylglucosamine (Merzendorfer and Zimoch 2003; Yang et al. 2008; Qiu et al. 2022). These enzymes are distributed in different organisms, since chitin is the second most present biopolymer in the world, participating in different physiological processes (Qiu et al. 2022). Chitinases have three primary functions in crustaceans: participating in the molting and growth processes of these animals, helping in the digestion of foods that contain chitin, and directly or indirectly participating in defense responses against bacteria, fungi, and viruses (Zhang et al. 2021). Although these molecules have the ability to kill some chitin-containing pathogens through their catalytic activity (Zarei et al. 2011; Seo et al. 2016), they seem to play a major role in direct and indirect immune regulation (Niu et al. 2018; Zhang et al. 2021). The LvChi5 chitinase from Penaeus vannamei does not show in vitro activity against Vibrio parahaemolyticus or Escherichia coli, but expression of LvChi5 was induced against V. parahaemolyticus, white spot syndrome virus (WSSV), lipopolysaccharides (LPS), and poly(I:C) (Niu et al. 2018).The absence of LvChi5 through silencing by RNAi caused an increase in the mortality of P. vannamei against infections with WSSV and V. parahaemolyticus. Moreover, silencing of this gene also resulted in transcriptional inhibition of immune genes such as antimicrobial peptides, transcriptional factors, and other proteins with antibacterial or antiviral activity (Niu et al. 2018). Chitinase transcriptional levels of WSSV-resistant Penaeus japonicus were higher than virus-sensitive animals (Pan et al. 2005). The three chitinases differently expressed found in the present study are encoded by different genes and share an aminoacidic identity ranging from 54.2% to 68.2% to each other. Chitinases are very diverse molecules, being divided into two families with multiple groups according to sequence homology and catalytic mechanism with multiple functions, which could be pathogen-specific or gene-specific regulations (Zhang et al. 2021). Therefore, the activity of some chitinases may be mainly due to immune regulation and these genes could play a major role in the ISD development through multiple routes.
As well as chitinases, B-GlcNacases comprise a diverse family of intracellular, secreted, or transmembrane molecules that appears to participate in immune responses (Zhang et al. 2021; Soto-Rodriguez et al. 2022). Gene expression of a GlcNacase from Exopalaemon carinicauda increased in the hepatopancreas after infection with V. parahaemolyticus and Aeromonas hydrophila (Sun et al. 2018). In mammals, GlcNacase is one of the preformed mediator lysosomal enzymes, which is stored in some granulocytes and is released upon infection (Prabhuling et al. 2021). These molecules are also associated with intercellular communication during immune responses in mammals (Prabhuling et al. 2021). In cell cultures of Drosophila melanogaster, silencing of a lysosomal GlcNacase by RNAi resulted in an increase of mycobacteria growth (Koo et al. 2008). In Bombyx mori, GlcNacase activity is increased during haemocytic degranulation after parasitism, indicating the release of this molecule to fight the parasite directly or indirectly (Prabhuling et al. 2021). The mucin-like protein is found in the membrane of the hepatopancreas and is associated with protection against bacteria (Soto-Rodriguez et al. 2022), but some contradictory studies suggest an importance of mucin-like protein for bacterial establishment (De los Santos et al. 2022). It would be important to understand the role of mucin-2-like protein during ISD in future studies, since this was the only chitin-related gene that was more expressed in symptomatic animals.
A2M protein is a class of protease inhibitors abundant in the haemolymph of invertebrates, and acts as a multifunctional protein (Chaikeeratisak et al. 2012). They act as protease inhibitors neutralizing pathogenic proteases important for bacterial establishment (Armstrong 2010). However, these molecules are also involved in other immune responses, such as phagocytosis, prophenoloxidase activating systems, and the haemolymph clotting system (Chaikeeratisak et al. 2012). The gene modulation of A2M in the hepatopancreas and haemocytes of crustaceans during bacterial and viral infections has been reported (Pongsomboon et al. 2011; Ren et al. 2019). Previous studies found that the A2M expression was higher in symptomatic animals with ESD (Tarrant et al. 2010), which is contrary to the findings of this study (Tarrant et al. 2010). This suggests that, although ISD and ESD are similar in appearance, some immune responses against these two infections may be different.
Trypsin-1-like was also more expressed in As animals compared to others, which may be indicative of an immune response against ISD pathogens. Trypsin is related to metabolic processes but may also contribute to defense mechanisms in crustaceans in the form of a serine protease. Interestingly, the gene expression of trypsin-1 is frequently upregulated during infections in crustaceans (Clark et al. 2013a, 2013b, 2013c; Li et al. 2018; Yu et al. 2022). Trypsin may also be associated with the cleavage of haemocyanin into haemocyanin-derived peptides with immune activity. The regulation of trypsin against infections is related to the generation of haemocyanin-derived peptides in P. vannamei and its silencing by RNAi reduces the amount of haemocyanin-derived peptide. In addition, in vitro experiments have shown that recombinant trypsin hydrolyzes haemocyanin and the generated fragments have bacterial agglutinating capacity (Li et al. 2018). In line with this idea, a pseudohaemocyanin gene was also more expressed in As animals compared to symptomatic animals in this study. Pseudohaemocyanin function is not well understood, and there is no in vitro study showing its cleavage to produce fragments with antimicrobial activity. However, there is evidence of this molecule being cleaved, and the possibility of pseudohaemocyanin-derived peptides with antimicrobial activity should not be discarded (Terwilliger et al. 2005). Therefore, it is possible that the increase in trypsin and pseudohaemocyanin results in an increase in the number of fragments with antimicrobial activity to prevent the establishment of the disease.
Acute phase serum amyloid A or SAA is a protein present in the haemolymph of crustaceans associated with the acute phase of infections and is used as a biomarker in the medical and veterinary fields (Cray et al. 2009). In this work, the SAA gene was more expressed in MS animals compared to the As group (FDR < 0.05), and this trend was also observed without statistical difference in the comparison between SS and As animals (FDR = 0.7). These results suggest that MS animals are at the acute stage of the disease, whereas SAA levels in SS animals have started to decline. This could also indicate that As animals are, in fact, in a healthy condition. Although SAA is classically a biomarker of the acute stage of a disease, this gene is also upregulated in moribund lobsters infected with the Gram-positive bacteria Aerococcus viridans var. homari and the ciliated protist Anophryoides haemophila (Clark et al. 2013a, 2013b). Altogether, it is possible to suggest that, despite having an advanced stage of ISD, the acute phase of the disease is over in SS animals.
Conclusion
This was the first study to apply massively parallel sequencing techniques and the genome of H. americanus to investigate genes associated with the presence of ISD. We identified that 134 genes are differentially expressed in symptomatic and As ISD animals. Interestingly, most of these genes are more expressed in As animals compared to symptomatic animals, which could indicate a potential susceptibility or immunocompromise of symptomatic animals or resistance of As animals. Among the DEGs found are eight chitin-related proteins, suggesting that chitin and its related proteins are important to fight this infection. A2M and SAA also appear to be involved in ISD, and SAA may be a potential marker for the early stages of this disease. Future work is required to better understand how chitin and chitin-related proteins are associated to ISD defense or disease pathogenicity. In addition, it is essential to understand the defense mechanisms of lobsters against ISD in other tissues, such as the carapace epithelium of these animals, which is directly linked to this disease.
Acknowledgements
We are grateful to Clark Lab members for helping with sample collection. We are also grateful for Ocean Frontier Institute, NSERC/CRSNG (RGPIN-2018-5894) and A. David Crowe Graduate Scholarship for funding this project and scholarship, Genome Québec for sequencing, and usegalaxy.eu and usegalaxy.org for bioinformatic tools.
References
Afgan E., Baker D., Batut B., Van Den Beek M., Bouvier D., Ech M., et al. 2018. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2018 update. Nucleic Acids Research, 46(1): W537–W544.
Altschul S.F., Madden T.L., Schäffer A.A., Zhang J., Zhang Z., Miller W., Lipman D.J. 1997. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Research, 25(17): 3389–3402.
Andrews S. 2010. FastQC a quality control tool for high throughput sequence data 2010. Available from http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ [accessed December 2023].
Armstrong P.B. 2010. Role of α 2-macroglobulin in the immune responses of invertebrates. Invertebrate Survival Journal, 7: 165–180.
Bonilla-Gómez J.L., Chiappa-Carrara X., Galindo C., Jeronimo G., Cuzon G., Gaxiola G. 2012. Physiological and biochemical changes of wild and cultivated juvenile pink shrimp farfantepenaeus duorarum (Crustacea: Penaeidae) during molt cycle. Journal of Crustacean Biology, 32(4): 597–606.
Cawthorn R.J. 2011. Diseases of American lobsters (Homarus americanus): a review. Journal of Invertebrate Pathology, 106(1): 71–78.
Chaikeeratisak V., Somboonwiwat K., Tassanakajon A. 2012. Shrimp alpha-2-macroglobulin prevents the bacterial escape by inhibiting fibrinolysis of blood clots. PLoS ONE, 7(10): e47384.
Clark K.F., Acorn A.R., Greenwood S.J. 2013a. Differential expression of American lobster (Homarus americanus) immune related genes during infection of Aerococcus viridans var. homari, the causative agent of Gaffkemia. Journal of Invertebrate Pathology, 112(2): 192–202.
Clark K.F., Acorn A.R., Greenwood S.J. 2013b. A transcriptomic analysis of American lobster (Homarus americanus) immune response during infection with the bumper car parasite Anophryoides haemophila. Developmental & Comparative Immunology, 40(2): 112–122.
Clark K.F., Greenwood S.J., Acorn A.R., Byrne P.J. 2013c. Molecular immune response of the American lobster (Homarus americanus) to the white spot syndrome virus. Journal of Invertebrate Pathology, 114(3): 298–308.
Cole J.N., Nizet V. 2016. Bacterial evasion of host antimicrobial peptide defenses. Microbiology Spectrum, 4(1):.
Cray C., Zaias J., Altman N.H. 2009. Acute phase response in animals: a review. Comparative Medicine, 59(6): 517–526. Available from http://www.ncbi.nlm.nih.gov/pubmed/20034426 [accessed December 2023].
Dai L., Xiong Z., Hou D., Wang Y., Li T., Long X., et al. 2022. Pathogenicity and transcriptome analysis of a strain of Vibrio owensii in Fenneropenaeus merguiensis. Fish & Shellfish Immunology, 130: 194–205.
de Lorgeril J., Saulnier D., Janech M.G., Gueguen Y., Bachère E. 2005. Identification of genes that are differentially expressed in hemocytes of the Pacific blue shrimp (Litopenaeus stylirostris) surviving an infection with Vibrio penaeicida. Physiological Genomics, 21(2): 174–183.
De los Santos M.V., Sánchez-Salgado J.L., Pereyra A., Zenteno E., Vibanco-Pérez N., Ramos-Clamont Montfort G., Soto-Rodriguez S.A. 2022. The Vibrio parahaemolyticus subunit toxin PirBvp recognizes glycoproteins on the epithelium of the Penaeus vannamei hepatopancreas. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 257: 110673.
Department of Fisheries and Oceans. 2022. Seafisheries landed value by province, 2021. Available from https://www.dfo-mpo.gc.ca/stats/commercial/land-debarq/sea-maritimes/s2021pv-eng.htm [accessed December 2023].
FAO. 2020. The state of world fisheries and aquaculture 2020. FAO.
Götz S., García-Gómez J.M., Terol J., Williams T.D., Nagaraj S.H., Nueda M.J., et al. 2008. High-throughput functional annotation and data mining with the Blast2GO suite. Nucleic Acids Research, 36(10): 3420–3435.
Kanehisa M., Furumichi M., Tanabe M., Sato Y., Morishima K. 2017. KEGG: new perspectives on genomes, pathways, diseases and drugs. Nucleic Acids Research, 45(1): D353–D361.
Kim D., Langmead B., Salzberg S.L. 2015. HISAT: a fast spliced aligner with low memory requirements. Nature Methods, 12(4): 357–360.
Klompmaker A.A., Chistoserdov A.Y., Felder D.L. 2016. Possible shell disease in 100 million-year-old crabs. Diseases of Aquatic Organisms, 119(2): 91–99.
Koo I.C., Ohol Y.M., Wu P., Morisaki J.H., Cox J.S., Brown E.J. 2008. Role for lysosomal enzyme β-hexosaminidase in the control of mycobacteria infection. Proceedings of the National Academy of Sciences, 105(2): 710–715.
Krueger F. 2012. Trim Galore 2012. Available from http://www.bioinformatics.babraham.ac.uk/projects/trim_galore/ [accessed December 2023].
Li C., Wang F., Aweya J.J., Yao D., Zheng Z., Huang H., et al. 2018. Trypsin of Litopenaeus vannamei is required for the generation of hemocyanin-derived peptides. Developmental & Comparative Immunology, 79: 95–104.
Li W., Wang Q. 2021. Recent progress in the research of exosomes and Dscam regulated crab antiviral immunity. Developmental & Comparative Immunology, 116: 103925.
Liao Y., Smyth G.K., Shi W. 2014. FeatureCounts: an efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics, 30(7): 923–930.
Love M.I., Huber W., Anders S. 2014. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biology, 15(12): 1–21.
Meres N.J., Ajuzie C.C., Sikaroodi M., Vemulapalli M., Shields J.D., Gillevet P.M. 2012. Dysbiosis in epizootic shell disease of the American lobster (Homarus americanus). Journal of Shellfish Research, 31(2): 463–472.
Merzendorfer H., Zimoch L. 2003. Chitin metabolism in insects: structure, function and regulation of chitin synthases and chitinases. Journal of Experimental Biology, 206(24): 4393–4412.
Moriya Y., Itoh M., Okuda S., Yoshizawa A.C., Kanehisa M. 2007. KAAS: an automatic genome annotation and pathway reconstruction server. Nucleic Acids Research, 35(SUPPL.2): W182–W185.
Niu S., Yang L., Zuo H., Zheng J., Weng S., He J., Xu X. 2018. A chitinase from pacific white shrimp Litopenaeus vannamei involved in immune regulation. Developmental & Comparative Immunology, 85: 161–169.
Pan D., He N., Yang Z., Liu H., Xu X. 2005. Differential gene expression profile in hepatopancreas of WSSV-resistant shrimp (Penaeus japonicus) by suppression subtractive hybridization. Developmental & Comparative Immunology, 29(2): 103–112.
Pertea M., Pertea G.M., Antonescu C.M., Chang T.C., Mendell J.T., Salzberg S.L. 2015. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nature Biotechnology, 33(3): 290–295.
Pethybridge S.J., Nelson S.C. 2015. Leaf Doctor: a new portable application for quantifying plant disease severity. Plant Disease, 99(10): 1310–1316.
Polinski J.M., Zimin A.V., Fraser Clark K., Kohn A.B., Sadowski N., Timp W., et al. 2021. The American lobster genome reveals insights on longevity, neural, and immune adaptations. Science Advances, 7: 8290–8313.
Pongsomboon S., Tang S., Boonda S., Aoki T., Hirono I., Tassanakajon A. 2011. A cDNA microarray approach for analyzing transcriptional changes in Penaeus monodon after infection by pathogens. Fish & Shellfish Immunology, 30(1): 439–446.
Prabhuling S.H., Makwana P., Pradeep A.N.R., Vijayan K., Mishra R.K. 2021. Release of mediator enzyme β-hexosaminidase and modulated gene expression accompany hemocyte degranulation in response to parasitism in the silkworm Bombyx mori. Biochemical Genetics, 59(4): 997–1017.
Qiu S., Zhou S., Tan Y., Feng J., Bai Y., He J., et al. 2022. Biodegradation and prospect of polysaccharide from crustaceans. Marine Drugs, 20(5): 310.
R Core Team. 2023. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Available from https://www.R-project.org/ [accessed 13 December 2023].
Ren X., Liu P., Li J. 2019. Comparative transcriptomic analysis of Marsupenaeus japonicus hepatopancreas in response to Vibrio parahaemolyticus and white spot syndrome virus. Fish & Shellfish Immunology, 87: 755–764.
Ridgway I.D., Taylor A.C., Atkinson R.J.A., Stentiford G.D., Chang E.S., Chang S.A., Neil D.M. 2006. Morbidity and mortality in Norway lobsters, Nephrops norvegicus: Physiological, immunological and pathological effects of aerial exposure. Journal of Experimental Marine Biology and Ecology, 328(2): 251–264.
Robinson M.D., McCarthy D.J., Smyth G.K. 2009. edgeR: a bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics, 26(1): 139–140.
Seo D.J., Lee Y.S., Kim K.Y., Jung W.J. 2016. Antifungal activity of chitinase obtained from Paenibacillus ehimensis MA2012 against conidial of Collectotrichum gloeosporioides in vitro. Microbial Pathogenesis, 96: 10–14.
Smolowitz R.M., Bullis R.A., Abt D.A. 1992. Pathologic cuticular changes of winter impoundment shell disease preceding and during intermolt in the American Lobster, Homarus americanus. The Biological Bulletin, 183: 1.
Soto-Rodriguez S.A., Lozano-Olvera R., Montfort G.R.C., Zenteno E., Sánchez-Salgado J.L., Vibanco-Pérez N., Aguilar Rendón K.G. 2022. New insights into the mechanism of action of PirAB from Vibrio parahaemolyticus. Toxins, 14(4): 243.
Sun Y., Zhang J., Xiang J. 2018. Molecular characterization and function of β-N-acetylglucosaminidase from ridgetail white prawn Exopalaemon carinicauda. Gene, 648: 12–20.
Sun Y.D., Fu L.D., Jia Y.P., Du X.J., Wang Q., Wang Y.H., et al. 2008. A hepatopancreas-specific C-type lectin from the Chinese shrimp Fenneropenaeus chinensis exhibits antimicrobial activity. Molecular Immunology, 45(2): 348–361.
Tang D., Chen M., Huang X., Zhang G., Zeng L., Zhang G., et al. 2023. SRplot: a free online platform for data visualization and graphing. PLoS ONE, 18(11 October): e0294236.
Tarrant A.M., Stegeman J.J., Verslycke T. 2010. Altered gene expression associated with epizootic shell disease in the American lobster, Homarus americanus. Fish & Shellfish Immunology, 29(6): 1003–1009.
Terwilliger N.B., Ryan M.C., Towle D. 2005. Evolution of novel functions: cryptocyanin helps build new exoskeleton in Cancer magister. Journal of Experimental Biology, 208(13): 2467–2474.
Theriault M., Vanleeuwen J., Morrison M., Cawthorn R. 2008a. Risk factors for the development of shell disease in impounded populations of the American lobster, Homarus americanus. Journal of Shellfish Research, 27(5): 1239–1245.
Theriault M., Vanleeuwen J., Morrison M., Cawthorn R. 2008b. Risk factors for the development of shell disease in impounded populations of the American lobster, Homarus americanus. Journal of Shellfish Research, 27(5): 1239–1245.
Tlusty M.F., Smolowitz R.M., Halvorson H.O., DeVito S.E. 2007. Host susceptibility hypothesis for shell disease in American lobsters. Journal of Aquatic Animal Health, 19(4): 215–225.
Vogt G. 2019. Functional cytology of the hepatopancreas of decapod crustaceans. Journal of Morphology, 280(9): 1405–1444.
Warnes G., Bolker B., Bonebakker L., Gentleman R., Huber W., Liaw A., et al. 2022. Gplots: Various R Programming Tools for Plotting Data 2022. Available from https://CRAN.R-project.org/package=gplots [accessed December 2023].
Yang Q., Liu T., Liu F., Qu M., Qian X. 2008. A novel β-N-acetyl-d-hexosaminidase from the insect Ostrinia furnacalis (Guenée). The FEBS Journal, 275(22): 5690–5702.
Yu D., Zhai Y., He P., Jia R. 2022. Comprehensive transcriptomic and metabolomic analysis of the Litopenaeus vannamei hepatopancreas after WSSV challenge. Frontiers in Immunology, 13.
Zarei M., Aminzadeh S., Zolgharnein H., Safahieh A., Daliri M., Noghabi K.A., et al. 2011. Characterization of a chitinase with antifungal activity from a native Serratia marcescens B4A. Brazilian Journal of Microbiology, 42(297): 1017–1029.
Zhang X., Yuan J., Li F., Xiang J. 2021. Chitin synthesis and degradation in crustaceans: a genomic view and application. Marine Drugs, 19(3): 153.
Supplementary material
Supplementary Material 1 (XLSX / 11 KB).
- Download
- 10.97 KB
Supplementary Material 2 (XLSX / 122 KB).
- Download
- 121.02 KB
Supplementary Material 3 (DOCX / 438 KB).
- Download
- 437.14 KB
Information & Authors
Information
Published In

FACETS
Volume 9 • Number 1 • January 2024
Pages: 1 - 10
Editor: Mark D. Fast
History
Received: 18 March 2024
Accepted: 9 July 2024
Version of record online: 28 October 2024
Notes
This paper is part of a collection entitled One Ocean Health.
Copyright
© 2024 The Author(s). This work is licensed under a Creative Commons Attribution 4.0 International License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Data Availability Statement
Data generated or analyzed during this study are available in the NCBI-SRA repository, Accession number PRJNA1084878, https://www.ncbi.nlm.nih.gov/sra/PRJNA1084878.
Key Words
Sections
Subjects
Plain Language Summary
Impoundment shell disease: a transcriptomic immune perspective about an ignored lobster disease
Authors
Author Contributions
Conceptualization: NA
Formal analysis: NA
Investigation: NA
Software: MR
Supervision: KFC
Writing – original draft: NA
Writing – review & editing: KFC
Competing Interests
The authors declare there are no competing interests.
Funding Information
Natural Sciences and Engineering Research Council of Canada: RGPIN-2018-5894
A David Crowe Scholarship
Ocean Frontier Institute: (One Ocean Health)
Metrics & Citations
Metrics
Other Metrics
Citations
Cite As
Nicolas Argenta, Marianna Rampaul, and K. Fraser Clark. 2024. American lobster (Homarus americanus) hepatopancreas transcriptome reveals the significance of chitin-related genes during impoundment shell disease. FACETS.
9: 1-10.
https://doi.org/10.1139/facets-2024-0049
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
There are no citations for this item