Long-term ocean and resource dynamics in a hotspot of climate change
Abstract
The abundance, distribution, and size of marine species are linked to temperature and nutrient regimes and are profoundly affected by humans through exploitation and climate change. Yet little is known about long-term historical links between ocean environmental changes and resource abundance to provide context for current and potential future trends and inform conservation and management. We synthesize >4000 years of climate and marine ecosystem dynamics in a Northwest Atlantic region currently undergoing rapid changes, the Gulf of Maine and Scotian Shelf. This period spans the late Holocene cooling and recent warming and includes both Indigenous and European influence. We compare environmental records from instrumental, sedimentary, coral, and mollusk archives with ecological records from fossils, archaeological, historical, and modern data, and integrate future model projections of environmental and ecosystem changes. This multidisciplinary synthesis provides insight into multiple reference points and shifting baselines of environmental and ecosystem conditions, and projects a near-future departure from natural climate variability in 2028 for the Scotian Shelf and 2034 for the Gulf of Maine. Our work helps advancing integrative end-to-end modeling to improve the predictive capacity of ecosystem forecasts with climate change. Our results can be used to adjust marine conservation strategies and network planning and adapt ecosystem-based management with climate change.
Graphical Abstract

Introduction
Marine ecosystems vary naturally across timescales, exhibiting seasonal, annual, decadal, centennial, and longer cycles. In addition, humans have gradually intensified their influence on marine ecosystems since the last ice age through exploitation, habitat alteration, and biogeochemical changes, thereby modifying species abundances and ecosystem structure resulting in intergenerational shifts of observational “baselines” (Pauly 1995; Jackson et al. 2001; Lotze and Worm 2009). The superposition of human impacts on natural long-term variability has made it difficult to understand the causes of some complex, nonlinear ecosystem changes including recent collapses of fish stocks and regime shifts (Lindegren et al. 2010; Auber et al. 2015; Pershing et al. 2015; Friedland et al. 2019, 2020) and poses challenges to marine conservation and resource management.
The Northwest Atlantic Ocean (NW Atlantic) has supported major fisheries and other marine resource uses over centuries (Lotze and Milewski 2004; Rose 2004; Rosenberg et al. 2005; Alexander et al. 2009) and continues to support significant fisheries and livelihoods for people. Two prominent marine ecoregions, the Gulf of Maine and Scotian Shelf, are located at the confluence of major northward (Gulf Stream) and southward (Labrador Current) ocean currents with strong influences on marine ecosystem dynamics (Fig. 1). The relative strength and position of these currents are sensitive to climate variability on interannual to multidecadal and longer time scales, including the North Atlantic Oscillation, Atlantic Multidecadal Oscillation, Atlantic Meridional Overturning Circulation, and anthropogenic climate change (Greene and Pershing 2000, 2003; Saba et al. 2016; Brickman et al. 2018; Caesar et al. 2018, 2021; Claret et al. 2018). These ecoregions could therefore serve as barometers for broader climate variability and ecosystem changes in the NW Atlantic and globally (Greene and Pershing 2000, 2003; Saba et al. 2016).
Fig. 1.

Currently, the NW Atlantic is experiencing rapid shifts in climate conditions and is considered a hotspot of climate change impacts (Caesar et al. 2018, 2021; Brickman et al. 2018; Thibodeau et al. 2018; Thornalley et al. 2018; Mellon et al. 2019). These changes will alter marine ecosystems and fisheries, as already observed in the decline of Gulf of Maine cod (Pershing et al. 2015), a reorganization of fish community structure (Friedland et al. 2019) and changes in lower trophic levels (Friedland et al. 2020). It is not always easy, however, to differentiate the effects of climate variability from those of fishing without a more thorough understanding of climate and ecological variability over longer time scales (Bograd et al. 2003; Rose 2004; Free et al. 2019). Despite the ecological and economic importance of the Gulf of Maine and Scotian Shelf, knowledge of premodern oceanographic and ecosystem baselines remains limited. Although paleoceanographic, archaeological, and historical records exist (e.g., Scott et al.1984; Lear 1998; Lotze and Milewski 2004; Rosenberg et al. 2005; Bourque et al. 2008; Betts et al. 2017), these have not been synthesized in a comprehensive way.
This paper reviews and synthesizes available paleoceanographic, archaeological, historical, and modern records and model projections from the Gulf of Maine and Scotian Shelf to acquire a long-term multidisciplinary perspective on ocean environmental and ecosystem changes. Our goals are to assess: (i) how the NW Atlantic has changed over the past >4000 years, (ii) how long-term environmental changes and human influences have affected resource abundance and ecosystem structure, and (iii) how future projections under climate change compare to past trends. Thereby, we identify critical gaps in record coverage, potential additional data sources, and uncertainties when combining records from multiple disciplines. We further evaluate the implications of our findings for advancing past-to-future marine ecosystem modeling and how our results can inform decision making and help adapt marine conservation strategies and ecosystem-based management with climate change.
Focus region
The focus region covers the Gulf of Maine from Cape Cod to the Scotian Shelf as far north as the Laurentian Channel (Fig. 1) and consists of shallow banks interspersed with >250 m deep ocean basins. This temperate NW Atlantic region is fed by three major water sources: the warm, salty Gulf Stream from the south; the cold, less saline Labrador Current from the north; and freshwater from the St. Lawrence River (Fig. 1). The Labrador Current consists of two southwestward flowing branches; the inshore branch flows into the Gulf of St. Lawrence, mixing with fresh St. Lawrence River water before exiting back through the Cabot Strait on to the Scotian Shelf, forming the Nova Scotia Current (Beardsley and Boicourt 1981; Han and Loder 2003; Dever et al. 2016). This Scotian Shelf water eventually makes its way into the Gulf of Maine, where it travels anti-cyclonically around the semi-enclosed basin, while the offshore branch of the Labrador Current moves southwestward along the slope of the Scotian Shelf break (Rutherford and Fennel 2018). Slope waters enter the deep basins of the shelf through several cross-shelf channels (Shan et al. 2016), carrying remineralized nutrients (Petrie and Yeats 2000; Townsend et al. 2015).
Although the Scotian Shelf and Gulf of Maine ecosystems are highly connected, they are separated by the international border between Canada and the United States (US). For our study, we focus on ocean dynamics on the continental shelf (Fig. 1) but also utilize fisheries and marine ecosystem data from the Exclusive Economic Zones (EEZs) and Northwest Atlantic Fisheries Organization (NAFO) management areas. We focus on the last ∼4000–5000 years, which include the late Holocene cooling and recent warming and thus major periods of climate change (Scott et al. 1984; Edgecombe et al. 1999; Levac 2001). Moreover, by ∼5000 years before present (BP) postglacial sea level rise had reached approximately −10 m (Vacchi et al. 2018) and slowed significantly thereafter; therefore, large-scale bathymetry and regional oceanography has not changed fundamentally since (Barnhardt et al. 1995; Shaw et al. 2002). This means that coastal archaeological sites cover this time span, as earlier settlement sites were eroded and drowned during the Holocene marine transgression (Shaw et al. 2009; Kelley et al. 2013). The earliest human presence likely occurred after the last Ice age ∼13,000 BP (Spiess et al. 1990; Betts and Hrynick 2021), with places such as the Turner Farm in Penobscot Bay (Bourque et al. 2008), Passamaquoddy Bay in the Outer Bay of Fundy (Lotze and Milewski 2002, 2004), and Port Joli on the Atlantic shore of Nova Scotia (Betts et al. 2011, 2017) (Fig. 1) occupied repeatedly over the past ∼4500, ∼4000, and ∼2000 years, respectively. This pre-Colonial period ended with the arrival of Europeans beginning in the ∼1500s (Lotze and Milewski 2004; Lotze 2010; Betts and Hrynick 2021). Since then, the region has experienced major changes in human population growth, land and coastal transformation, technology and industrialization, and fishing intensity (Lotze and Milewski 2004; Rosenberg et al. 2005; Bourque et al. 2008; Lotze 2010).
To synthesize long-term climate, environmental, and ecological changes, we considered data from paleoceanography, archaeology, history, and modern ocean sciences. First, we compiled records of climate and other environmental changes from “archives” in sediment cores, deep-water corals, mollusk shells, historical and modern instrumental records, and model simulations (Figs. 1 and 2). Next, we compiled available information on resource and ecosystem changes based on records of fish and other marine vertebrates from microfossils in sediment cores, archaeological shell middens, historical records and reconstructions, and modern monitoring data. We also compiled future projections of climate, environmental, and ecosystem changes from available model projections. The following sections are structured according to these data sources.
Fig. 2.

Climate and water temperature changes
High-resolution in situ time series measurements are generally limited in spatial and temporal extent. Instrumental records on multidecadal to centennial scales rely on few coastal monitoring stations or ship-based observations (Deser et al. 2010), some extending back to the early 19th century (Freeman et al. 2017). Our study region is close to major shipping lanes, and it is possible to reconstruct temperature variations on multidecadal time scales, including estimates of uncertainty based on space–time density of available observations. In contrast, paleoclimate archives in sediments can reveal patterns over hundreds or thousands of years. In the following, we synthesize available monitoring data with paleoceanographic proxy records and global climate model simulations to reconstruct sea surface temperature (SST) over the last 4000 years and project future trends until 2100. We also calculate the time of emergence, the year when the mean SST anomaly surpasses its long-term natural variability.
Temperature records
Estimates of SST were drawn from (i) global gridded ship- and buoy-based products, (ii) coastal monitoring stations, (iii) paleoceanographic proxies, and (iv) global climate models.
Five global gridded in situ monthly SST data sets consisted of two types (Table S1): (i) ERSST v5 (Huang et al. 2017), HadISST v1 (Rayner et al. 2003), and COBE SST2 (Hirahara et al. 2014) used statistical methods to interpolate SST data over space and time and don’t contain missing data fields; and (ii) CERA-20C (Laloyaux et al. 2018) and SODA (Giese et al. 2016) are reanalysis products that assimilate diverse climate observations into a numerical ocean model. The spatial resolution of data sets ranged from 0.5–2.0°, and the time periods cover starting dates from January 1854 to December 1901 and end between December 2010 and July 2019 (Table S1).
Three in situ monthly SST data series from coastal monitoring stations in Halifax and St. Andrews (Fig. 1, Table S1) were derived from Fisheries and Oceans (DFO) Canada (Petrie and Drinkwater 1993) and in Boothbay Harbor from the Maine Department of Marine Resources (Drinkwater and Petrie 2011). Measurements have been taken twice daily and averaged monthly since the early 20th century from near-surface water bucket samples until July 1993, which were then replaced with moored digital temperature recorders at 2 m depth. The percent of missing data is 5.1% at Halifax, 2.2% at St. Andrews, and 0.6% at Boothbay.
Continuous paleoceanographic SST reconstructions with multidecadal resolution were available from two sediment cores (OCE326-MC29, OCE326-GGC30) from the Scotian Shelf (Keigwin et al. 2003; Sachs 2007) and one (OCE400-MC44) from Jordan Basin, Gulf of Maine (Keigwin and Pilskaln 2015; for revised age models see Mellon et al. 2019) (Figs 1 and 2a, Table S1). SSTs from OCE326-MC29 and OCE326-GGC30 were estimated from alkenone paleothermometry using the Prahl et al. (1988) temperature calibration, which measures the molecular make-up (unsaturation ratio) of alkenones (long-chain carbon molecules) produced by planktonic coccolithophores and reflects the organism’s growth temperature (Brassel 1986). Alkenone paleothermometry has been a robust proxy of SST in the NW Atlantic, except in regions with permanent winter sea ice (Filippova et al. 2016). SST from OCE400-MC44 (Gulf of Maine) was determined from stable oxygen isotope ratios (δ18O) of planktonic foraminifera. Keigwin and Pilskaln (2015) demonstrated that δ18O of Neogloboquadrina incompta is a significant linear function of SST based on modern proxy calibrations in the Gulf of Maine.
Climate model data were derived from the global CESM1 model, specifically the Large ENSemble (LENS) set of experiments (Kay et al. 2015). The model produces SST estimates from 1920–2100 using a fully coupled CMIP5 global climate simulation with 40 ensemble members. The model output consists of three different experiments produced with different forcings. From 1920–2005, the “historical” experiment was subjected to natural (solar, volcanic) and anthropogenic radiative forcing. From 2006 onward, the RCP8.5 experiment is forced by projected future anthropogenic greenhouse gas emissions under the worst emissions and low mitigation scenario (Riahi et al. 2011). We also examined the 1800 year “control” (or “natural”) experiment which is subjected to pre-industrial levels of radiative forcing (Table S1). The CESM-LENS does not include any other emissions scenarios (Kay et al. 2015), such as the strong mitigation scenario RCP2.6; however, emissions since 2005 and the likely trajectory of near-future emissions closely follow the RCP8.5 scenario until at least 2040 (Pedersen et al. 2020).
Temperature trajectories
Piecing these records together, we calculated SST anomalies (SSTA) relative to a baseline period of 1920–1950, which was shared by all temperature records. We also derived the natural mean (and percentiles) from the global climate model natural control run (Fig. 3). Lastly, we calculated centennial-scale slopes in SSTA because the temporal resolution between past, current, and future records highly varied (Fig. 4). This was done by determining the linear regression slope of the SSTA over a 100-year moving window.
Fig. 3.
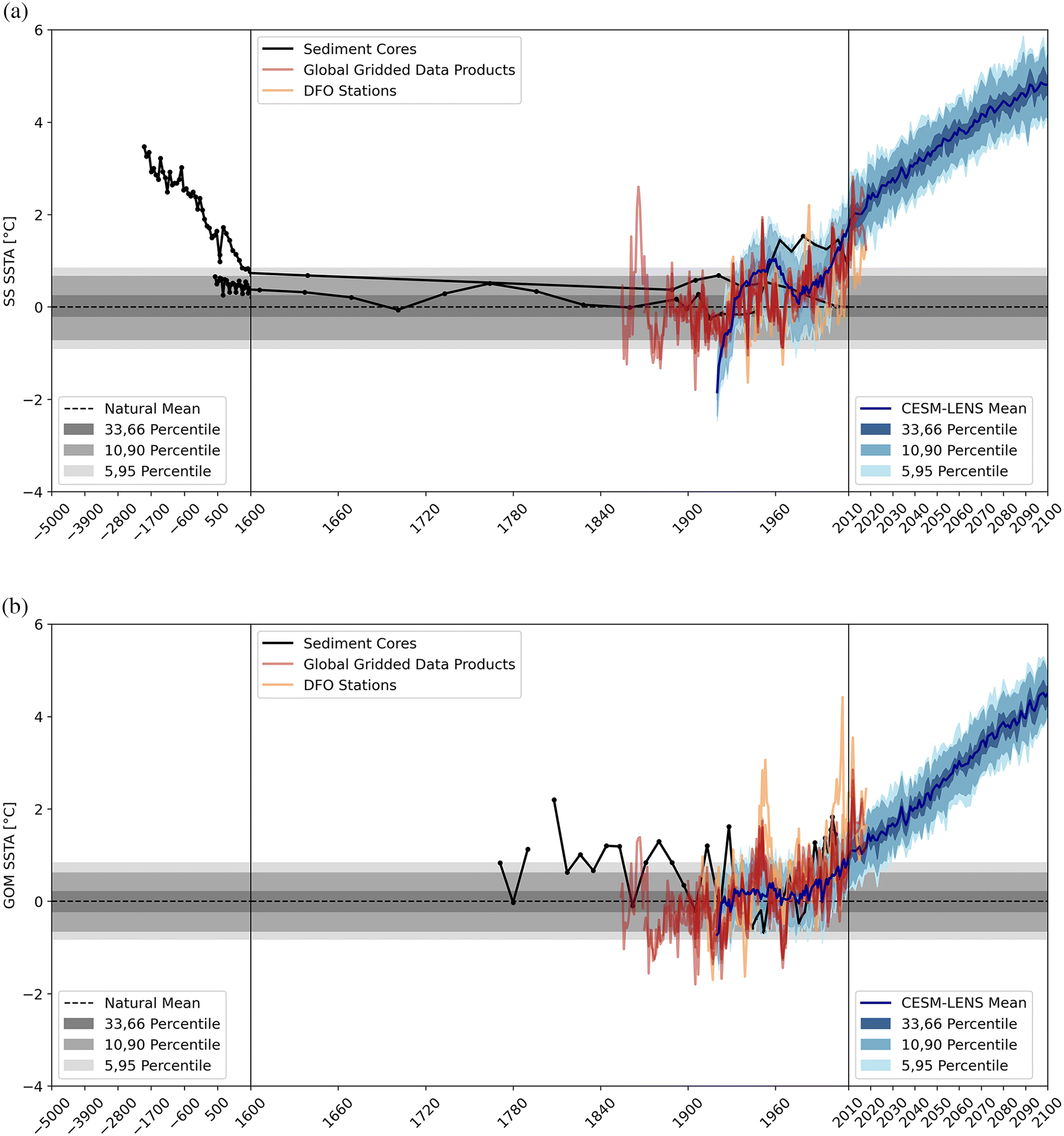
Fig. 4.

Compared to the baseline or natural mean, the alkenone-based SST reconstructions from the longer core OCE326-GGC30 suggest a ∼2.5 °C cooling from 1800 BC to 1800 AD on the Scotian Shelf (Fig. 3a; Sachs 2007), consistent with an overall Holocene cooling of the NW Atlantic (Leduc et al. 2010). The shorter core OCE326-MC29 did not exhibit this signal (Keigwin et al. 2003), likely because the core is not long enough to record this trend (see Mellon et al. 2019 for uncertainties in age model). Other proxy records also imply relative cooling in the NW Atlantic during the Holocene (Scott et al. 1984; Keigwin and Jones 1995; Marchitto and deMenocal 2003). For example, dinoflagellate assemblages from 10 500–8 000 BP in La Have Basin, Scotian Shelf, reflect ∼5 °C higher SST than present (Levac 2001; see Mircofossils section), and a 1–2 °C cooling over the last 1000 years has been found in mollusk shell archives from the Gulf of Maine (Wanamaker et al. 2008; see Mollusk archives section). Amplified Holocene cooling along the NW Atlantic shelf has been attributed to AMOC variability, specifically a more southerly position of the Gulf Stream and intensified Labrador Current influence (Levac 2001; Sachs 2007; Wanamaker et al. 2008). Not all proxy records show this Holocene cooling in the NW Atlantic (e.g., Mg/Ca ratios of planktic foraminifera in Keigwin et al. 2005); however, this is likely due to different seasonal and depth preferences of different proxy signal carriers, which is the focus of ongoing debate (e.g., Leduc et al. 2010).
More recent SSTAs show multidecadal oscillations along with a steady increase since ∼1960 (Fig. 3), most noticeable in coastal monitoring stations and global gridded data. On the Scotian Shelf, these two data sources show a ∼0.5 °C warming around 1950 and 1970 with a cooling in between (Fig. 3a) and a short warming period in one of the global gridded data products around 1860. Similar multidecadal oscillations are seen in the Gulf of Maine with a prominent warming around 1950 followed by a cooling around 1960 in both the coastal monitoring stations and global gridded data (Fig. 3b). The global climate model data also show a strong multidecadal oscillation on the Scotian Shelf between 1920–2010, which is not mirrored in the Gulf of Maine, and a large negative tail at the time series start.
The majority of SSTAs from coastal monitoring stations and global gridded data between 1850 and 2010 are within the bounds of natural SSTA variability from the CESM-LENS natural control run (Fig. 3). On the Scotian Shelf, 77.6%–92.4% of global gridded data SSTAs are within the 5,95-percentile natural window from the start of each time series to 2010; in the Gulf of Maine this amounts to 70.7%–91.7%. The Halifax coastal monitoring station has 79.7% of SSTAs within the 5,95 percentile bounds, compared to 73.0% for St. Andrews and 52.9% for Boothbay.
Since ∼1960, coastal monitoring stations, global gridded data, and climate models all show a clear warming trend in both regions (Fig. 3) and CESM-LENS projects consistent warming from 2005–2100. By 2100, SSTA is projected to reach +4.8 °C on the Scotian Shelf and +4.5 °C in the Gulf of Maine. These projections are in line with regional, higher-resolution simulations (Brickman et al. 2021; Rutherford 2021; see General circulation models section).
On the Scotian Shelf, the future SSTA increase is projected to have a comparable amplitude to the decrease prior to 1600 seen in alkenone-based SSTA reconstructions (Fig. 3a). Importantly, the cooling reconstructed from sediment cores occurred over ∼3500 years while the projected warming occurs over just 150 years; this difference is apparent when analyzing the warming rate (Fig. 4). Due to the large time window in which the cooling takes place, the sediment core-derived SSTA slope is consistently within the bounds of natural variability, while the projected CESM-LENS mean SSTA is consistently outside. This holds for the Gulf of Maine and Scotian Shelf and accounts for ensemble variability (Fig. 4). There is a noticeable difference during 2010-2050; however, the Scotian Shelf SSTA slope plateaus and then decreases whereas the Gulf of Maine SSTA slope decreases in magnitude but remains positive. Most SSTA slopes from global gridded data also surpass the window of natural variability.
Time of emergence
Using the natural control run along with historical and future CESM-LENS runs allows determining the time of emergence (ToE), defined as the year when SSTA consistently exceeds 95% of the natural variability for the rest of the time series (Henson et al. 2017). This can be interpreted as the point at which the climate change signal emerges from the range of historical natural variability. First, a year is selected when the climate change signal begins by determining when the cumulative sum of the SSTA gradient exceeds 0 for the rest of the time series. A linear slope (w) is determined from this selected year to the end of the time series. The standard deviation (σ) of the CESM-LENS natural control run is also determined, explaining 65% of total natural SSTA variability under assumptions of normality; σ is multiplied by 2 to have bounds at 95% of natural variability, assuming normality. The ToE is then calculated as:
For natural climate variability, each of the data products should not surpass the 95% percentile bands determined from the CESM-LENS natural control run. All global gridded data time series eventually surpass this natural window of variability after 2010 (Fig. 3). Assuming that the natural SSTA variability for CESM-LENS matches the natural variability in the global gridded data, then this emergence can be attributed to anthropogenic forcing. The resulting ToE for the CESM-LENS mean SSTA would be 2028 on the Scotian Shelf and 2034 in the Gulf of Maine, as expected by the sharp increase in SSTAs in both regions.
Ocean environmental changes
In addition to temperature, several other environmental factors have changed over time, but have generally less resolved long-term trends. In the following, we provide an overview of other data sources that can be used to understand long-term environmental changes: (i) deep-water corals, (ii) mollusk archives, (iii) monitoring data, and (iv) general circulation models.
Deep-water corals
Deep-water corals represent paleoceanographic archives of high longevity and temporal resolution (Robinson et al. 2014; Williams 2020). Certain gorgonian corals found along the NW Atlantic margin generate growth rings in their skeletons (Fig. 2b) over several hundreds of years (Sherwood and Edinger 2009). The skeletons contain a diagenetically resistant, fibrillar protein derived from sinking organic matter originating in surface waters (Sherwood et al. 2006). Geochemical analysis of the growth rings provides annually resolved, centennial-length proxy records of surface water processes. For example, stable nitrogen isotope ratios (δ15 N) in growth rings of Primnoa resedaeformis from the Northeast Channel (Fig. 1) can be interpreted as a proxy for nitrate source (accounting for trophic level and other effects on δ15 N) and hence the relative influence of Gulf Stream or Warm Slope Water versus Labrador Current or Labrador Slope Water contributing to export productivity (Sherwood et al. 2011). These records show that colder Labrador Current water dominated since at least 400 AD, whereas the intrusion of warmer Gulf Stream water since the early 1970s is a relatively recent phenomenon. These results are in agreement with SST reconstructions based on other lines of evidence (Fig. 3), but they also show that the variable mixture of different water masses in our study region affects physical and biochemical (e.g., nutrient) conditions (see Monitoring data section).
Mollusk archives
Marine bivalve shells show growth lines (Fig. 2d) that can be analyzed with high-resolution isotope sclerochronology that aligns daily, monthly, or annual shell growth to oxygen isotope records. Bivalves from archaeological shell middens (Fig. 2e) or marine sediments are thus valuable proxies for reconstructing environmental records of past sea surface conditions (Wanamaker et al. 2007; Burchell et al. 2014). Using palaeothermometry for calcite or aragonite (Grossman and Ku 1986), shell-derived oxygen isotope data (δ18O) can be converted to temperature. Historical and modern ocean quahog (Arctica islandica) shells from the western Gulf of Maine showed a mean SST of 8.5 (±1) °C during ∼1033-1062 AD, 7.5 (±1) °C during ∼1321–1391 AD, and 6.4 (±0.8) °C in the 20th century, suggesting a 1–2 °C cooling over the past ∼1000 years (Wanamaker et al. 2008). The quahog shells further showed a 1.6 °C or δ21% reduction in seasonal temperature range in ∼1033–1062 AD compared to ∼1321–1391 AD and 1864–1886 AD, as well as colder summers and warmer winters in the 11th century compared to the 14th and late-19th century (Wanamaker et al. 2011). This decreased seasonality possibly resulted from increased stratification of coastal waters due to warmer temperatures, whereas cooling during the 14th and 19th centuries was associated with increased vertical mixing that enhanced the seasonal temperature amplitude. Preliminary SST reconstructions from δ18O of Mya arenaria shells from Port Joli dating ∼600–1700 AD revealed a range of ∼8–22 °C with an average minimum of ∼12 °C and average maximum of ∼18 °C. In comparison, modern SST ranges from 0.0 °C in February to 18.8 °C in September (DFO 2021a).
Archaeological shells can also be used to study intra- and inter-shell variability in radiocarbon signals that affect calibrations and SST reconstructions (Dusseault 2021). In conjunction with oxygen isotope analysis, radiocarbon dates can also be used to study variation in coastal upwelling (Monge and Alveirinho 2006; Ferguson et al. 2013; Etayo-Cadavid et al. 2019) or to calculate a geographically localized marine reservoir correction (ΔR) necessary for calibrating marine radiocarbon dates (Betts 2019; Heaton et al. 2020). Combining the season of death with an approximate age of harvested bivalves further permits long-term interpretation of the frequency, intensity, and season of shellfish harvesting and thus insight into cultural practices (Burchell et al. 2014; Ambrose et al. 2016; Betts et al. 2017).
Monitoring data
Several physical and chemical water-column properties have been monitored over past decades (Bernier et al. 2018; NOAA 2019, 2021a). In addition to temperature, this includes information on ocean currents, stratification, sea level, oxygen, acidification and nutrients. For example, the Atlantic Zone Monitoring Program (AZMP) has employed a network of sampling locations, including fixed high-frequency sampling stations, cross-shelf sections, and ecosystem trawl surveys that have been sampled twice-monthly to once-annually since 1998. Long-term changes in ocean properties have been summarized in Bernier et al. (2018), in annual reports by DFO (e.g., Casault et al. 2020; Hebert et al. 2021), and in biannual reports by the National Oceanographic and Atmospheric Administration (e.g., NOAA 2019, 2021a).
Increasing air temperatures (∼1 °C per century since the 1870s) and the growing influence of the Gulf Stream relative to the Labrador Current are leading to warmer sea surface and deep-water temperatures in the Gulf of Maine and Scotian Shelf (Bernier et al. 2018; NOAA 2021a). Warming waters have extended the summer season, with a ∼2-week earlier arrival of the spring thermal transition date and a ∼2-week later arrival of the fall thermal transition date, affecting species’ phenology such as the timing of the spring bloom (NOAA 2019, 2021a). Since the late 19th century, gauging stations in Halifax and Saint John, New Brunswick, have recorded a 2–4 mm per year sea level rise (Bernier et al. 2018). Warming waters together with higher freshwater inputs to surface waters have enhanced water column stratification since 1948, reducing vertical mixing (Bernier et al. 2018). Carbon dioxide concentrations causing ocean acidification have increased more strongly in Canada’s Atlantic waters than other parts of the world; the pH on the Scotian Shelf has decreased by ∼0.03 units per decade on average (Bernier et al. 2018). There has also been a decline in oxygen content, particularly in deeper waters (Bernier et al. 2018). Changes in nutrient conditions have been more variable; however, nitrate concentrations generally declined from 1999 to 2010 (Bernier et al. 2018). Silicate concentrations have also decreased in the North Atlantic due to a weakening of the subpolar gyre, weaker vertical mixing, and stronger influence of subtropical surface water (Hátún et al. 2017).
The interplay of subarctic and subtropical current systems in our study region has strong influences on temperature and salinity as well as nutrient and oxygen distributions because of different biogeochemical processing histories. Labrador Current waters originate, in part, from Pacific waters that travel through the North American Arctic as a subsurface halocline layer before reaching the northern Labrador Shelf (Tremblay et al. 2015). High productivity on the shelves and upper slopes of the Bering and Chukchi seas fuels high rates of sedimentary denitrification, imparting a characteristic nitrate deficit to the Pacific water (Yamamoto-Kawai et al. 2006). Large river systems draining most of the Canadian landmass also contribute to an excess of silicate and phosphate. The resulting nutrient anomalies (nitrate concentration <15 μM; nitrate:silicate <1:1, nitrate:phosphate <16:1) in this colder, fresher water can be traced along the length of the Labrador Current into the Gulf of Maine and the mid-Atlantic Bight (Jones et al. 2003; Townsend et al. 2010; Fennel 2010). Conversely, Warm Slope Water (also known as Atlantic Temperate Slope Water) originates from the Sargasso Sea thermocline. Remineralization of organic matter in these deeper, warmer, saltier waters leads to higher nitrate concentrations (>23 μM) and Redfield-like nitrate:silicate (1.1:1) and nitrate:phosphate (16:1) ratios (Jenkins et al. 2015). The remineralization also leads to a characteristic depletion in oxygen saturations (<50%). In both cases, however, the deeper slope waters that enter the shelf areas through deep channels are more nutrient rich than shallower shelf and surface waters (Fig. 1) and become available to the euphotic zone through vertical mixing or upwelling. The relative contribution of these different water masses and the water column stratification determine the prevailing regime of biochemical conditions (Townsend et al. 2010).
During the 1960s cold period, when Labrador Slope Water dominated deep (>150 m) inflows to the basins of the Scotian Shelf and Gulf of Maine, nitrate concentrations, nitrate:silicate and nitrate:phosphate ratios were at lower levels and oxygen saturation at higher levels than at any point in the instrumental record (Petrie and Yeats 2000; Townsend et al. 2010). Coral- and bivalve-based δ15 N records, which serve as a proxy for Pacific water (Sherwood et al. 2021), indicate that this colder, lower-nitrate regime likely persisted over most of the post-European settlement period (Sherwood et al. 2011; Whitney et al. 2019), generally consistent with the SSTA records in Fig. 3. In the early 1970s, Warm Slope Water began to dominate the on-shelf flow with corresponding increases in nitrate concentrations, nitrate:silicate and nitrate:phosphate ratios and decreases in oxygen saturations. This trend to warmer waters with more Atlantic-like nutrient and oxygen distributions has continued to present (Claret et al. 2018; Casault et al. 2020). However, increasing water column stratification limits the availability of these deeper slope-water nutrients to shallower shelf and surface waters, resulting in a projected decrease in nitrate concentrations on the shelves (see General circulation models section).
General circulation models
General circulation models (GCMs) simulate physical and chemical properties and circulation patterns of the atmosphere and oceans and can be used to project climate change impacts (Figs. 5 and 6). Generally, GCM results for the NW Atlantic are consistent with a high sensitivity of the region to climate change. Comparisons of available observations with the few existing high-resolution model simulations suggest major changes in circulation are underway and will accelerate under strong warming (Brickman et al. 2021; Rutherford 2021). Lower-resolution global models typically used for climate projections struggle to capture regional dynamics (Loder et al. 2015; Lavoie et al. 2017; Laurent et al. 2021), leading to uncertainty in projections.
Fig. 5.

Fig. 6.

Regional, higher-resolution simulations yield significantly improved representations of local hydrography and biogeochemical dynamics compared to global models of the Coupled Model Intercomparison Project Phase 5 (CMIP5) and 6 (CMIP6) (Rutherford and Fennel 2018; Brickman et al. 2021; Laurent et al. 2021). A major contributor to the lower skill of global models is the small spatial scale of essential circulation features of the Labrador Current System (Fig. 1), including the shelf-break current that effectively isolates shelf water from the adjacent open ocean resulting in long shelf residence times and distinct water properties (Rutherford and Fennel 2018). The shelf break current is a relatively narrow jet that cannot be represented in coarser global models, yet its absence results in vastly different shelf residence times (Rutherford and Fennel 2018) and contributes to the discrepancies in biogeochemical properties compared to observations (Laurent et al. 2021). But, although high-resolution regional models resolve local hydrography well, future projections require inputs from the atmosphere and along their boundaries, which can only be derived from global models.
The global Geophysical Fluid Dynamics Laboratory (GFDL) model CM2.6 provides a unique combination of high resolution and global scale. This model includes a fully coupled ocean and atmosphere and simulates a weakening of the AMOC related to a dramatic warming on the NW Atlantic shelf in response to increasing CO2 (Saba et al. 2016). The simulated warming is accompanied by declining dissolved O2 along the upper slope; both changes are related to decreasing influx of cold, oxygenated Labrador Sea water and increasing dominance of warm Gulf Stream water (Claret et al. 2018). We used the 10-km regional biogeochemical model (Laurent et al. 2021) to downscale a CO2-doubling simulation of CM2.6 for the time slice 2065–2080. The time axis of the CM2.6 simulation was stretched to resemble the medium emission scenario RCP6.0 (Claret et al. 2018). In the future time slice, the shelf break current weakens substantially along the Grand Banks and northeastern Scotian Shelf and vanishes along the southwestern Scotian Shelf (Figs. 5a, 5b). This leads to relatively little warming and freshening of bottom waters on the Grand Banks while the southwestern Scotian Shelf and, to a lesser extent, the Gulf of Maine become saltier and warmer (Figs. 5c–f) due to inflow of slope waters with a high proportion of Gulf Stream water (Rutherford 2021). Nutrient supply on the Scotian Shelf and Gulf of Maine generally decreases in the future time slice (Figs. 6a, 6b) as the nutricline moves downward with increasing stratification. The fastest rate of bottom-water acidification of 0.3 pH units occurs on the Grand Banks compared to 0.1 pH units on the southwestern Scotian Shelf and Gulf of Maine (Figs. 6c, 6d). The smaller pH decline in the Gulf of Maine and southwestern Scotian Shelf is linked to increased inflow of warm, salty slope water and decreased delivery of subpolar waters (Salisbury and Jönnson 2018). In contrast, bottom-water O2 concentrations are projected to slightly increase on the Grand Banks but decrease by >40 mmol·m−3 on the southwestern Scotian Shelf and ∼20 mmol·m−3 in the Gulf of Maine (Figs. 6e, 6f), another consequence of inflow of less ventilated Gulf Stream water.
Focusing on the Gulf of Maine, Brickman et al. (2021) used two high-resolution numerical ocean models to downscale global climate projections and derive four estimates of physical properties in 2050 under business-as-usual emission scenarios. All simulations project increases in mean surface (1.1–2.4 °C) and bottom (1.5–2.1 °C) temperatures and increased water column stratification. Salinity changes are more variable among simulations, but mostly decrease in surface and increase in bottom waters (Brickman et al. 2021).
The projected future changes in physical and chemical properties are in line with environmental monitoring trends and may allow informed inferences about past changes of these parameters during the Holocene cooling (Fig. 3). This would mean that with cooling, lower water-column stratification, and increased influence of the Labrador Current relative to the Gulf Stream, waters on the Scotian Shelf and Gulf of Maine would have become less saline, more nutrient-enriched, more oxygenated, and less acidic over at least the past 1000 years (Fig. 3). The higher silicate:nitrate ratios in Labrador-sourced waters would have favoured subarctic, larger-cell diatom-based primary productivity (Harrison and Li 2008; Townsend et al. 2010). These trends would have had a positive influence on marine production.
Primary production and plankton communities
Changes in climate and ocean environment directly or indirectly influence organisms on all levels of the food web (Worm and Lotze 2021). In this section we synthesize changes in net primary production, phytoplankton, and zooplankton communities based on: (i) microfossils, (ii) monitoring data, and (iii) available future projections.
Microfossils
Most plankton species do not have any hard parts that are preserved in the sedimentary record, except a few groups that leave shells, cysts, organic linings, or other structural parts that can provide a snapshot into ancient plankton communities. For example, a sediment core from Wilkinson Basin, Gulf of Maine, revealed different phases in diatom species composition since the last glacial period, and the diatom flora approached modern Gulf of Maine conditions by ∼5,000 BP with increasing dominance of Thalassiosira eccentrica and associated species (Schnitker and Jorgensen 1990). Using a multi-proxy approach to determine changes in diatoms, dinoflagellates, foraminifera, and pollen spores in sediments from Bedford Basin in Halifax, Scotian Shelf Miller et al. (1982) showed clear assemblage changes alongside a transition from freshwater to estuarine to fully marine conditions during the early-mid Holocene. An increase in sedimentation rate over the past 250 years coincided with increased land clearing, run-off, and sewage discharges and was associated with a two-fold decrease in dinocyst and benthic foraminifera abundance (Miller et al. 1982).
Using a high-resolution sediment-core record from La Have Basin, Scotian Shelf, Levac (2001) reconstructed sea surface conditions over the past 10 000 years and found a succession of major paleoceanographic events and associated changes in dinoflagellate cysts (dinocysts) concentrations and assemblage composition. The dinocyst assemblage showed highest productivity, including blooms of toxic, red-tide producing algae (e.g., Alexandrium excavatum) in the early Holocene >7500 BP, when waters were warmer and saltier, with possibly nutrient-rich conditions and greater water column stability. In contrast, dinocyst concentrations were lower during the colder, less saline late Holocene period. The highest production of pelagic foraminifera followed the dinocyst peak, while the highest production of benthic foraminifera was more in line with the dinocyst peak suggesting that increased marine productivity occurred over the entire water column (Levac 2001). A similar record of peak abundance of red-tide producing dinocysts in the warmer, early Holocene was found in Emerald Basin (Mudie et al. 2002), while the late Holocene cooling was associated with increased predominance of arctic dinocysts (Scott et al. 1984). In both La Have and Emerald Basin, harmful algal blooms coincided with 2-5 °C warmer summer SSTs, while they were much less prevalent during later, colder periods. Similar observations on Canada’s Pacific coast indicate that climate warming may be a major driver in stimulating harmful algal blooms in the past and future (Mudie et al. 2002).
Monitoring data
Warming waters together with changes in other physical and chemical parameters are causing shifts in phytoplankton and zooplankton abundance and changes in assemblage composition and phenology as observed in long-term monitoring programs (Bernier et al. 2018; NOAA 2019, 2021a; Casault et al. 2020) and other studies (e.g., Townsend et al. 2010; Pershing et al. 2021).
Both the AZMP and NOAA found gradual declines in chlorophyll-a and phytoplankton abundance in the NW Atlantic since 1999, suggesting a reduction in production potential (Bernier et al. 2018; NOAA 2019, 2021a). This is supported by a study of available chlorophyll-a measurements over the past century indicating a 100-year decline in chlorophyll-a concentrations in NW Atlantic (Boyce et al. 2010). Generally, increasing water column stratification restricts the inflow of nutrient-rich deeper waters through vertical mixing and upwelling thereby limiting productivity levels, and these trends are projected to increase in the future (Fig. 6b, see Future projections section). In addition, the ratios of different nutrients are important drivers for phytoplankton composition. A long-term comparison of scientific plankton surveys from the 1930s and 1990s in the Outer Bay of Fundy showed an overall reduction of the spring phytoplankton bloom and a shift in species composition with a decrease in diatoms and increase in dinoflagellates (Lotze and Milewski 2002, 2004), a common consequence of nitrogen enrichment and eutrophication in inshore waters. In the Gulf of Maine, the increasing influence of warmer, saltier Warm Slope Water since the 1970s with higher nitrate has favoured a productivity regime with more dinoflagellates and warm-water plankton species (see Microfossils section), while the influence of colder, fresher Labrador Current water, with lower nitrate but higher silicate:nitrate and phosphate:nitrate ratios, has favoured a more subarctic productivity regime with large diatoms in the 1960s and before (Townsend et al. 2010; Pershing et al. 2021). Observed decreases in silicate concentrations in the North Atlantic will enhance silicate limitation of diatom blooms and lead to shifts in phytoplankton community composition, particularly shifts towards lightly silicified diatoms and nonsilicifying plankton (Hátún et al. 2017). Together, oceanographic conditions and species composition influence the timing, magnitude, and duration of the spring bloom, which showed trends toward an earlier start, a higher peak magnitude, and shorter duration from 1999 to 2010 (Bernier et al. 2018).
Observed shifts in phytoplankton abundance, composition, and phenology affect higher trophic levels (Bernier et al. 2018). Since 1999, changes have been observed in zooplankton composition suggesting important shifts at the food-web base. Generally, large, energy-rich Calanus finmarchicus copepods, which have been characteristic of subarctic plankton communities and a direct link between large diatoms and consumers in the past, have declined in abundance on the Scotian Shelf and Gulf of Maine, while smaller, warm-water copepods such as Pseudocalanus spp. and noncopepod zooplankton have increased (Bernier et al. 2018; Pershing et al. 2021). Other important zooplankton prey including euphausiids and shrimp have also shifted northward (Pershing et al. 2021). Such changes in food availability and composition affect zooplanktivorous consumers, such as forage fish (e.g., Atlantic herring (Clupea harengus) and sand lance (Ammodytes dubius), larval Atlantic cod (Gadus morhua), and the endangered North Atlantic right whale (Eubalaena glacialis, Pershing et al. 2021).
Future projections
A variety of model approaches exist to project future trajectories of net primary production (NPP), phytoplankton, and zooplankton biomass under different climate change scenarios. The Fisheries and Marine Ecosystems Model Intercomparison Project (Fish-MIP, isimip.org/about/marine-ecosystems-fisheries/, Tittensor et al. 2018; Lotze et al. 2019), for which we report changes in marine animal biomass below, used output of two global Earth System Models (ESMs) from CMIP5 (GFDL-ESM2M, IPSL-CM5ALR) that span the range of outcomes from other CMIP5 models. For both ESMs, Bryndum-Buchholz et al. (2020) derived outputs for NPP for the Gulf of Maine (NAFO 5YZe) and Scotian Shelf (NAFO 4XWV) under two opposing climate change scenarios, the currently observed high-warming scenario (RCP8.5) and a strong-mitigation scenario (RCP2.6). The ensemble means under RCP2.6 indicate a 10% and 11% increase in NPP by the 2030s and a slightly lower 6% and 7% increase by the 2090s on the Scotian Shelf and Gulf of Maine, respectively (Bryndum-Buchholz et al. 2020). In contrast, under RCP8.5, NPP was projected to decrease by −9% and −11% by the 2030s and by −13.5% and −13.7% by the 2090s in the Gulf of Maine and Scotian Shelf, respectively (Bryndum-Buchholz et al. 2020). This could be explained by the projected decreases in nitrate concentrations and increased stratification from high-resolution regional models due to warming (Fig. 6b).
Projected decreases in NPP are generally accompanied by slightly stronger decreases in phytoplankton biomass and even stronger decreases in zooplankton and higher trophic level biomass as the effect of reduced energy travels up the food chain (Lotze et al. 2019), a process called trophic amplification (Kwiatkowski et al. 2019). We can therefore expect similar changes with warming waters and lowered NPP in our study region. We note that coarse global ESMs often fail to integrate local- or regional-scale processes, particularly in coastal areas, which can alter fine-scale patterns and processes and thus projected trajectories (see above, Laurent et al. 2021), although the general trends of declining nitrate and NPP on the Scotian Shelf and Gulf of Maine seem to match. As with GCMs, regional or regionally downscaled global biogeochemical models could provide more refined future trajectories.
In the Gulf of Maine, several model approaches have simulated future abundance and distribution of the key zooplankton Calanus finmarchicus (Pershing et al. 2021). Reygondeau and Beaugrand (2011) projected that C. finmarchicus would disappear from the Gulf of Maine by 2050, based on a statistical model built on surface conditions, while Grieve et al. (2017) projected a ∼50% decline by end-century when including bottom water conditions. Using a life-history model, Ross et al. (2021) suggested a decline in summer and fall. While varying in the magnitude of change, all models agree on a general decline of C. finmarchicus.
Long-term changes in marine resources and ecosystems
Environmental conditions have direct (e.g., oxygen, temperature) effects on consumers and indirect effects through altered prey and predator abundance. However, the relationship between long-term environmental conditions and fish or marine mammal populations is poorly constrained. Moreover, humans have used and influenced marine resources in the NW Atlantic for thousands of years, adding complexities to ecosystem dynamics. Several data sources exist to determine changes in resource abundance over past centuries and millennia and into the future (Lotze et al. 2011, Lotze and McClenachan 2014), including: (i) microfossils, (ii) archaeological records, (iii) historical records and hindcasting, (iv) modern scientific data, and (v) future projections. In the following, our focus is on records of biomass, relative abundance, or size rather than fisheries catches which are influenced by management decisions.
Microfossils
Fish teeth and scales have often been overlooked as paleoecological proxies in sediment cores due to their small size. Previous work in the California Current utilized fish scales from sediment cores to reconstruct shifts between sardine and anchovy populations over the past 2000 years partly related to environmental cycles (Baumgartner 1992; Finney et al. 2010). Fish otoliths from the Santa Barbara Basin revealed that mesopelagic fish biomass fluctuated with century-scale temperature variability (Jones and Checkley 2019), and fish teeth from a 3000-year sediment core from Panama suggested that parrotfish declines were linked to centuries of overfishing and recent coral reef degradation (Cramer et al. 2017). These windows into pre-industrial conditions provide baselines for understanding long-term ecosystem dynamics.
Few comparable studies have been undertaken in the NW Atlantic. However, ichthyoliths (fish teeth and shark scales; Sibert et al. 2017) extracted from sediment cores from Emerald Basin (Core 88010, 239.6 m depth) and the Scotian Slope (Core 2016011, 2360 m; Fig. 1, Table S2) revealed nearly an order of magnitude more fish remains in the 1800s compared to today and differences in assemblage composition (Fig. 2c). These preliminary results are in line with historical records and hindcasting results (see below) and suggest that ichthyoliths represent promising archives to reconstruct fish abundance, distribution, and community composition over time. Moreover, as samples from offshore shelf regions, they may reflect long-term changes in exploited and nontarget species and changes with environmental and anthropogenic conditions that could complement trends derived from archaeological records.
Archaeological records
Archaeofauna from shell middens along the coasts provide insights into the variety, size and relative abundance of marine species used by Indigenous people over time (Bourque et al. 2008; Betts et al. 2011, 2014, 2017; Lotze and Milewski 2004). After consumption, shells, bones, and other inedible parts were discarded on garbage piles that, over time, developed into mounds known as “shell middens”. Some shell middens have well-preserved layers that represent archives of biological, environmental, and human records. Earliest records go back ∼4500 years at the Turner Farm in Penobscot Bay, Gulf of Maine (Bourque et al. 2008), ∼4000 years in Passamaquoddy Bay, Outer Bay of Fundy (Lotze and Milewski 2002, 2004), and ∼1600 years in Port Joli, Nova Scotia (Betts et al. 2011, 2017) (Figs. 1 and 2e). This includes records on Atlantic cod and other marine fish, shellfish, and mammals (Table S2).
Several studies reconstructed cod size and age using archaeofauna (Rojo 1986, 1987, 1990; Kenchington and Kenchington 1993, Jackson et al. 2001; Betts et al. 2011, 2014) to provide pre-industrial baselines. Results suggest that people utilized large cod (>70–100 cm) for most of history, whereas average size of modern cod reached only 20–30 cm in coastal Gulf of Maine fisheries surveys and 40–60 cm in research and fisheries surveys in the Gulf of Maine and Scotian Shelf (Fig. 7), including several surveys utilizing fishing gear (e.g., jigs, handlines) comparable to historical methods (Jackson et al. 2001; Betts et al. 2011). This indicates an overall reduction in cod size and age largely attributed to overharvesting after European arrival, particularly with the rise of industrial fishing (Jackson et al. 2001; Betts et al. 2011). In the early data, slight increases in average cod size at Turner Farm and Port Joli coincided with Late Holocene cooling (Fig. 3) and were likely climate-driven (Betts et al. 2011).
Fig. 7.
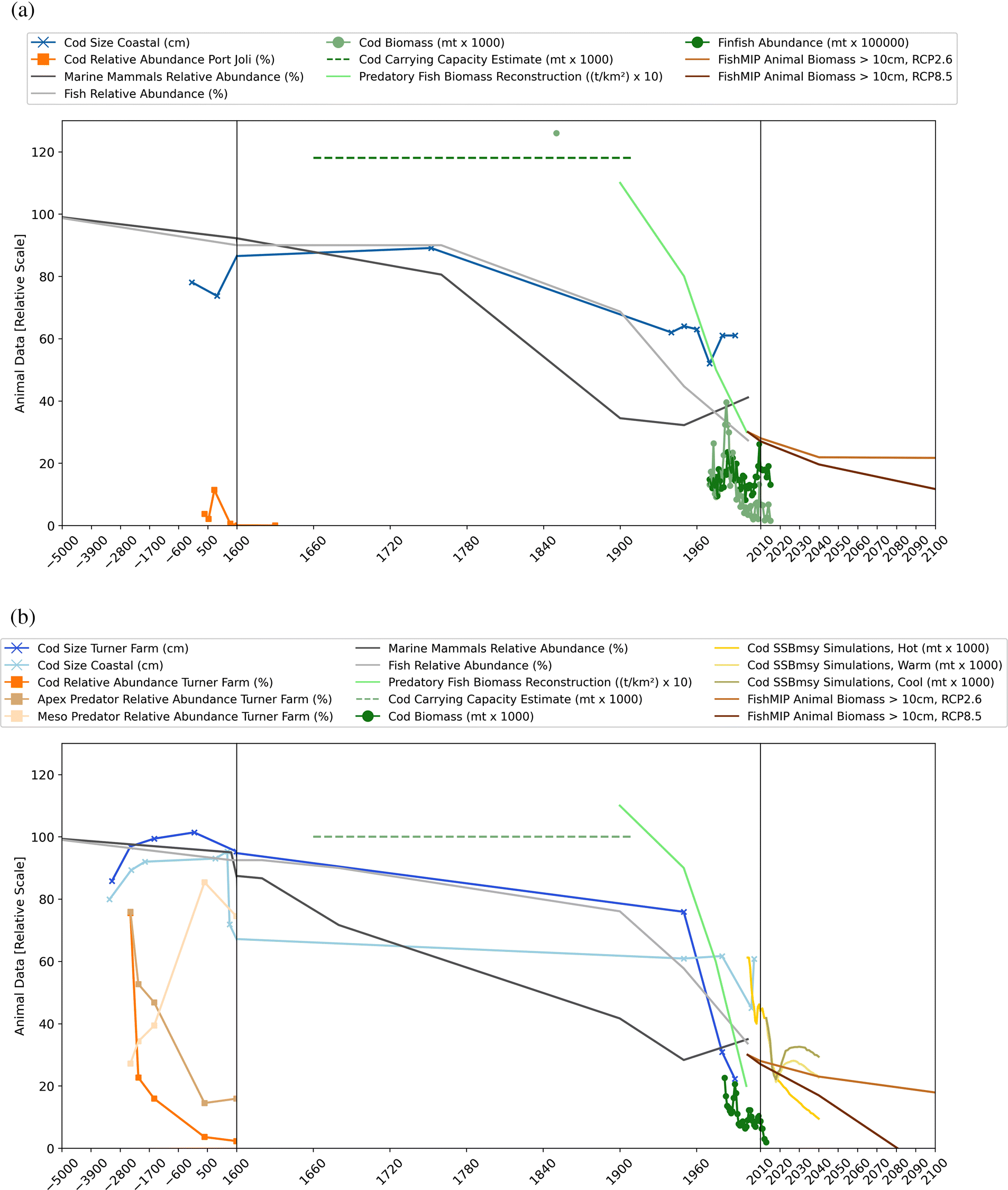
Archaeofauna analyses also include trends in relative abundance of cod and other marine species over time compared to other faunal remains. Results suggest an early decline of cod relative to other fish and vertebrate species at the Turner Farm site (Fig. 7b; Bourque et al. 2008), and a similar but much less pronounced trend at the Port Joli site (Fig. 7a; Betts et al. 2017). At the Turner Farm, there was also a steep early decline in the relative abundance of apex predators (including cod, swordfish, seals, sea mink, and dogfish) and an increase in mesopredators (including flounders, sculpins, tomcod, and other fish species) (Fig. 7b), whereas bivalves showed no trend over time (Bourque et al. 2008). These records suggest that at some sites, Indigenous fishing may have reduced local resource abundance, perhaps akin to microhabitat depression, although this was likely localized and limited to inshore areas (Bourque et al. 2008). With European arrival and technological progress, fishing expanded into offshore areas with sailing schooners in the 19th century and steamboats and otter trawls in the 20th century, thus increasing in spatial scale over time (Steneck 1997; Lotze and Milewski 2004). The early decrease in cod and apex-predators and increase in mesopredators could also stem from a diversification of the species spectrum fished, rather than overexploitation. Another possible explanation would be a shift in inshore oceanographic conditions resulting in resource depression, although the Late Holocene cooling seemed to positively affect average cod size due to colder, more productive waters (Betts et al. 2011). A general issue with relative abundance data from shell middens is that we cannot derive absolute abundance trends. Here, analysis of microfossil fish teeth and scales from sediment cores in offshore, less human-influenced areas could shed light on actual abundance trends in earlier times.
Historical records and hindcasting
For the historical period, available records include written descriptions by European naturalists or local fisheries inspectors and entries from fishers’ logbooks and sales or tax records that reach back to the 1500s and provide some insight into resource abundance. Generally, European explorers encountered plentiful and diverse resources along the coasts, and often mentioned large sizes (Lotze and Milewski 2002, 2004; Lotze 2010). Since the 1860s, there are also detailed annual catch statistics for commercially important species, while research survey and monitoring data provide observed abundance trends since the 1960s (Lotze and Milewski 2004; Alexander et al. 2009). Some of these records have been combined to reconstruct a cod biomass of ∼1.26 million metric tons (mt) on the Scotian Shelf in the 1850s (Rosenberg et al. 2005) compared to <400,000 mt on average in research surveys in the 1970s and <40,000 mt in the 1990s (Fig. 7), a > 95-fold decline mainly attributed to overfishing. Similarly, carrying capacity of cod on the Scotian Shelf was estimated at ∼1.18 million mt (Rosenberg et al. 2005), with a ∼95% reduction of cod biomass on the Scotian Shelf and similar values in the Gulf of Maine towards the end of the 20th century (Myers and Worm 2005). In the Gulf of Maine, annual fisheries landings of cod in the 1860s were estimated between 62,600–78,600 mt, >10 times higher than average landings towards the end of the 20th century (Alexander et al. 2009).
Historical reconstructions based on ecosystem models for predatory fish biomass also reveal strong declines of >70% and >80% from 1900-2000 for the Scotian Shelf and Gulf of Maine, respectively (Fig. 7) due to overfishing (Christensen et al. 2003). Another hindcasting approach used qualitative and quantitative archaeological, historical, and modern records to reconstruct relative abundance trends of ecologically and commercially important species in Massachusetts Bay, Gulf of Maine, and the Outer Bay of Fundy, Scotian Shelf (Lotze et al. 2006; Lotze 2010). This approach assumed pristine abundance (100%) of resource populations in prehuman times and, from there, traced relative declines and recoveries over time (Lotze et al. 2006). For marine mammals and fish, resulting trends revealed abundant populations (>90%–100%) during indigenous occupation over past millennia (Fig. 7), with only some depletions in highly valued and utilized species. This changed rapidly after European settlement with growing demand, commercialization of resource use, marketing of fashion and luxury products, and technological progress leading to enhanced effort, efficiency, and destructiveness of harvesting. Consequently, resource populations showed strong declines in abundance between 1600 and 1900 (Fig. 7). In the 20th century, depletion trajectories continued for fish, while marine mammals showed lesser declines in the first and some recoveries in the second half of the 20th century (Fig. 7), reflecting increased management and conservation (Lotze et al. 2006; Lotze 2010). By year 2000, the composite relative abundance compared to the pristine baseline reached 35%–41% for marine mammals and 27%–34% for fish in both regions (Fig. 7), although some species remain at much lower population levels or have gone extinct (Lotze and Milewski 2004; Lotze 2010). Interestingly, in all reviewed cases of species extinctions and depletions up to year 2000, climate change was never mentioned as a major driving factor (Lotze et al. 2006), although climate variability has always influenced population performance. Instead, overexploitation was the predominant cause of depletion and extinction followed by habitat loss, whereas the ban or restriction of exploitation and habitat protection were major drivers of recovery, and often both were needed to ensure success (Lotze et al. 2006).
Modern scientific data
Since the 1970s, standardized research surveys exist for groundfish communities in our study regions. On the Scotian Shelf, these surveys revealed declines in average cod biomass from 146 000 mt in the 1970s to 53 000 mt in 2005–2015 (Fig. 7), while overall finfish biomass has been more variable over time (Fig. 7) (Bundy et al. 2017; Bernier et al. 2018). In the Gulf of Maine, average cod biomass declined from 15,000 mt in the 1980s to 7 000 mt in 2003–2013 (Pershing et al. 2015). Importantly, cod biomass continued to decline despite a fishing moratorium since 1992. Several factors may explain the nonrecovery, including current populations being very depleted compared to pre-exploitation levels or carrying capacity (see Historical records and hindcasting section; Myers and Worm 2005; Rosenberg et al. 2005), overfishing of local subpopulations, continued cod bycatch in other fisheries, shifts in community composition, consumption by predators, changed oceanographic conditions (Myers et al. 1997; Rose 2004; Pedersen et al. 2017; Carson et al. 2017; Bernier et al. 2018) and increasingly climate change (see Future projections section, Pershing et al. 2015). This is corroborated by an analysis of changes in recruitment capacity in global fish stocks (Britten et al. 2016), which highlights an average 3% decline in recruitment over past decades worldwide, but higher declines in the North Atlantic, including the Scotian Shelf and Gulf of Maine, particularly for species like Atlantic cod. These declines were linked to historical overfishing, warming waters, and changes in primary production (Britten et al. 2016). A different approach hindcasted temperature-driven changes in maximum sustainable yield (MSY) of fish stocks worldwide (Free at al. 2019) and found an average 4.1% decrease in MSY from 1930 to 2010 due to warming. Different regions and species showed various responses, but generally Atlantic cod and other Gadiformes were negatively influenced by warming, although some fish stocks in the NW Atlantic showed positive responses (Free at al. 2019).
Shifts in individual species abundance and distribution affect community structure and food webs. In the Outer Bay of Fundy, overexploitation of traditionally common cod, haddock, and pollock in the 1990s led to strong increases in dogfish and hake (Lotze and Milewski 2004). Such compensation effects partly explain the more variable overall finfish biomass over past decades compared to strong declines in cod (Fig. 7) (Bundy et al. 2017). The overexploitation of predators can also result in increased abundance of forage fish, such as herring, which can cause a regime shift (Frank et al. 2005, 2011; Pedersen et al. 2017). Since the 1980s, however, strong declines in herring have been observed on the Scotian Shelf (Bernier et al. 2018). Marine invertebrates are also affected by the depletion of predators, with observed increases in lobsters, crabs, shrimp, sea urchins, and sea cucumbers in the Outer Bay of Fundy and on the Scotian Shelf over past decades (Lotze and Milewski 2004; Anderson et al. 2008).
Recent shifts in species composition have been linked to climate change, with cold-loving species moving further north and warm-loving species entering the Gulf of Maine and Scotian Shelf from the south (Pinsky et al. 2013; Bernier et al. 2018). On the Northeast US Shelf, 48 of the more abundant species have shifted their distribution northeastward and into deeper water (NOAA 2019). The decline in cod, warming waters, and influx of smaller fish species from the south has resulted in higher diversity and productivity in the fish community, but not necessarily overall biomass, a trend described as tropicalization (Friedland et al. 2019). These shifts reconfigure local and regional fish communities with often unpredictable results (Friedland et al. 2019, 2020; Lotze et al. 2021). In addition to the effects of warming, several species will also be affected by reduced oxygen concentrations (e.g., Atlantic wolffish; Bianucci et al. 2015) or ocean acidification (e.g., lobster; Waller et al. 2017).
Future projections
To gain insight into potential future trajectories of cod, fish, or marine animal biomass, several studies have employed model projections under different climate change scenarios. For cod in the Gulf of Maine, Pershing et al. (2015) used simulations of spawning stock biomass at their MSY (SSBmsy), which is temperature dependent. SSBmsy was ∼49 000 mt in 2000–2010 which declined to ∼12 022 mt under the currently hot climate change scenario in 2030–2040 compared to ∼25 092 mt under a warm and ∼31 254 under a cool scenario (Fig. 7), highlighting the benefits of climate change mitigation. Although these results have been contested by Brander (2018); several other studies employed species distribution projections for the Gulf of Maine and Scotian Shelf and all indicate that warming waters will lead to continued declines in thermal habitat area and biomass of key species, including Atlantic cod, haddock, and herring, while warm-adapted species from the south will migrate northward and increase (Shackell et al. 2014; Kleisner et al. 2017; Allyn et al. 2020; Pershing et al. 2021).
Bryndum-Buchholz et al. (2020) used simulations of marine animal biomass >10 cm (i.e., commercial size) from Fish-MIP under the observed high-warming scenario (RCP8.5) and a strong-mitigation scenario (RCP2.6) based on CMIP5. Results show continuing declines of animal biomass in the Gulf of Maine (NAFO 5YZe) reaching –12% under RCP2.6 and −38% under RCP8.5 by 2100 relative to the 1990s, compared to –8% and –18%, respectively, on the Scotian Shelf (NAFO 4XWV; Fig. 7). These simulations highlight the impact of climate change in the absence of fishing or other human impacts. We note, however, that fishing pressure will continue to play a major role in determining the status of current and future fish stocks and should be included in future modeling efforts, as well as the potential interaction between the effects of warming and fishing (Galbraith et al. 2017; Lotze et al. 2019).
Linking environmental and ecosystem changes
The late Holocene cooling over the past ∼4000 years (Fig. 3) likely had a positive effect on primary production and the size and abundance of cold-loving cod and other marine animals that have supported fisheries and other resource uses over millennia. This is reflected in slight increases in average size of cod in archaeological remains (Fig. 7) and the predominance of subarctic diatoms and copepods in the plankton community that may have favoured short, direct, and efficient food chains supporting higher trophic levels. The rate of temperature change during the late Holocene, however, was very low and not comparable to the recent rapid rate of warming (Fig. 4), allowing species plenty of time for acclimatization or adaptation. Until European arrival, most trends in marine fish and animal abundance and size were relatively stable (Fig. 7), suggesting that Indigenous hunting and fishing did not have major effects on overall resource abundance, except for some local decreases at Turner Farm (Fig. 7b; Bouruqe et al. 2008) and to a smaller extent at Port Joli (Fig. 7a; Betts et al. 2017). These trends suggest some localized resource depression in occupied inshore areas that were driven by either overexploitation, a diversification of resource use, localized climate variability, or a combination of these factors (Lotze et al. 2006; Betts et al. 2011, 2014).
Since European arrival, climate conditions were relatively stable until the later 20th century (Fig. 3). Therefore, most declines seen in cod and other marine animals between 1600–2000 (Fig. 7) can be attributed to overharvesting, which intensified in effort and efficiency, expanded in spatial scale, and diversified in resource use over time (Lotze and Milewski 2002, 2004; Rosenberg et al. 2005; Alexander et al. 2009; Lotze 2010) like in many other areas around the world (Jackson et al. 2001; Lotze et al. 2006).
Since the ∼1960s, a clear warming trend began (Fig. 3) with a rate of change not seen over the past ∼4000 years (Fig. 4). This recent warming, together with associated changes in other oceanographic conditions, comes on top of already high exploitation pressure and probably contributed to strong declines in cod and other cold-loving fish and marine species over past decades (Pershing et al. 2015; Britten et al. 2016; Free et al. 2019). The combined effects likely also contributed to the collapse of cod and other groundfish stocks in the early 1990s (Myers et al. 1997; Rose 2004; Carson et al. 2017; Pedersen et al. 2017). Because fishing pressure can increase the vulnerability of fish populations to climate change (Brander 2007; Lindegren and Brander 2018), the combined effects of climate change and (over)fishing can cause sudden shifts in population performance and fish stock collapses (Pershing et al. 2015).
Over the 21st century, climate change is projected to continue with warming waters (Fig. 3), declines in primary production, phyto- and zooplankton biomass, and associated decreases in cod and marine animal biomass (Fig. 7; Pershing et al. 2015; Bryndum-Buchholz et al. 2020). Cold-loving species will continue to shift towards colder waters further north, deeper, or offshore while warm-loving species will move in from the south (Shackell et al. 2014; Jones and Cheung 2015; NOAA 2019), resulting in marine communities and ocean ecosystems being reorganized in ways that have not occurred over the past ∼4000 years and are hard to predict (Friedland et al. 2019, 2020; Lotze et al. 2021; Pershing et al. 2021). The rapidity of warming may challenge species’ abilities to adapt or migrate, particularly less mobile or sessile species.
Although the current magnitude of warming mirrors SST conditions ∼1000–2000 BP, and by mid-century we may reach SSTs last seen ∼3000–4000 BP (Fig. 3), based on the available data there have been no obvious major changes in relative abundance, composition, or size of harvested fish or mammals between 4000–1000 BP that were explained by climate change, except some slightly enhanced marine production and size in harvested cod with Holocene cooling (Fig. 7) (Lotze and Milewski 2004; Rose 2004; Bourque et al. 2008; Lotze 2010; Betts et al. 2011, 2017). The changes in relative species composition in archaeological remains likely reflect localized resource depression, not a consequence of cooling, which would have favoured and not suppressed marine production. Interestingly, even when past temperatures were 2–4 °C warmer than in the late 20th century, it does not look like major species composition was dramatically different from today (e.g., presence of large cod and many other species still harvested today), while currently such a magnitude of warming is leading to the collapse of Gulf of Maine cod and pronounced range shifts in many species (Shackell et al. 2014; Pershing et al. 2015, 2021; NOAA 2019). There are few possible explanations for this; either the much slower rate of climate change may have allowed species to adapt and acclimatize to earlier warmer periods, or the much higher pre-industrial exploitation abundance made them more resilient to change or forced organisms to occur under suboptimal conditions because other habitats or niches were already occupied. In turn, today’s much lower abundance of exploited species may allow organisms to respond and move rapidly because habitat or niche space is available, or they are more vulnerable and sensitive to the combined effects of overfishing, climate change, and other human impacts. Looking ahead, current projections indicate a magnitude of warming by end-century that has last occurred >5000 BP in the early-mid Holocene warm period, when temperatures were about ∼5 °C warmer and harmful algal blooms were much more pronounced than over the past 4000 years (e.g., Levac 2001, Mudie et al. 2002). Currently, we do not know what the higher trophic level species composition would have been like >5000 BP, as most archaeological sites from that period have been submerged by sea level rise and no sediment cores with microfossil fish scales or teeth from that period have been recovered.
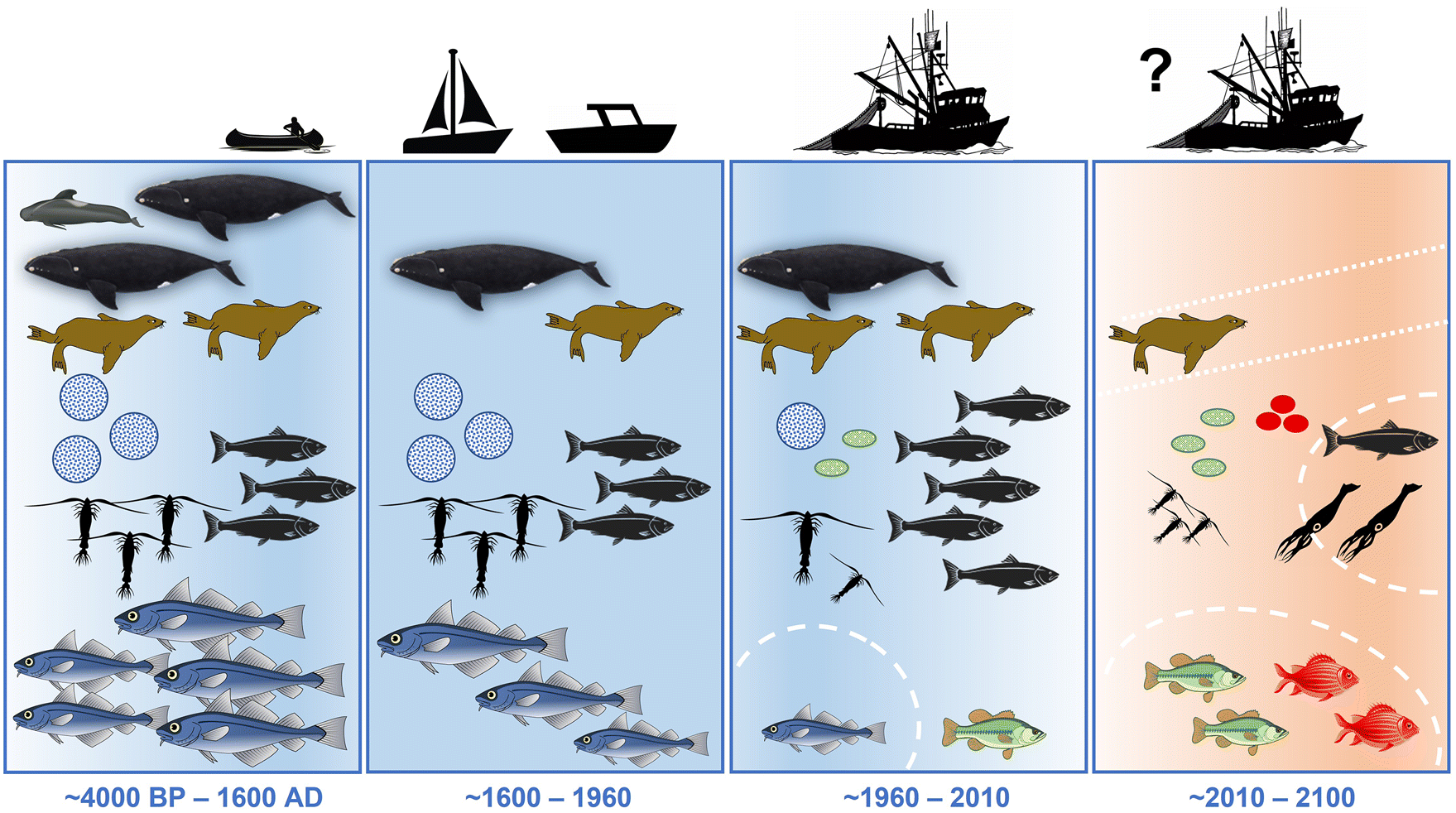
What can we learn from this? Based on the available evidence, the influence of climate and fishing/exploitation in the Gulf of Maine and Scotian Shelf ecosystem occurred in four major phases (Fig. 8):
1.
Influenced by colder, more productive subarctic waters, the late Holocene cooling probably had a positive impact on local resource populations that outweighed the fishing/exploitation effect by Indigenous people.
2.
Since European arrival up to the end-20th century, an increasingly intensifying fishing/exploitation effect caused strong depletion of many populations, while a relatively stable climate supported this history of exploitation (possibly similar to the stable land climate during the Holocene enabling the evolution of agriculture).
3.
Recent climate change came on top of already strong fishing pressure and other human impacts, thereby creating a double negative effect on already depleted populations (e.g., Gulf of Maine cod collapse) and shifts in fish community and ecosystem composition.
4.
Future projections indicate an increasingly negative climate effect with warmer, more stratified and less productive waters, particularly under a high-warming emission scenario (RCP8.5). By mid- to end-century, we are approaching climate and ecosystem conditions that have not occurred over the past ∼4000 years and are highly uncertain.
Fig. 8.
Implications for marine conservation, management, and policy
Our synthesis integrates multiple data sources across timescales to provide a long-term and multi-layered (physical-biological-anthropogenic) perspective on the Scotian Shelf and Gulf of Maine ecosystem with implications for marine conservation, management and policy. After relatively stable climate and ecosystem conditions over the past ∼4000-5000 years, recent rapidly changing ocean conditions and projected future warming and times of emergence emphasize the urgency to strengthen climate change adaptations in conservation, management, and policy if we are to sustain marine populations, ecosystem structure and functioning, fisheries, and human-dependent communities throughout the 21st century.
To counteract projected climate change impacts, a conservation and management process that takes a climate-adaptive, ecosystem-based, and precautionary approach is essential (Bryndum-Buchholz et al. 2021). This approach needs to account for shifting historical and future baselines, adjust exploitation rates and management goals, and develop climate-informed reference points that ensure populations and biodiversity remain at viable levels (Busch et al. 2016; Galbraith et al. 2017; DFO 2019a). This requires continued monitoring of changes in species productivity, abundance, and distribution. In the dynamic climate-change era, corresponding flexible and adaptive management tools are needed to respond to rapidly changing conditions (Dunn et al. 2016; McClenachan et al. 2019a; Bryndum-Buchholz et al. 2021). Currently, few Canadian fish stocks are assessed and managed considering climate change (DFO 2019b; Pepin et al. 2020). Vulnerability assessments provide an approach to triage species that are climate sensitive and require climate change to be incorporated into stock assessments (Stortini et al. 2015; Greenan et al. 2019). Climate-induced distribution shifts will have to be addressed both within and among nations through effective bilateral cooperation between scientists and managers (VanderZwaag et al. 2017). Within nations, fishing quotas and effort are often allocated based on management areas; the situation is further complicated for shifting transboundary species which nations assess and manage separately (Maureaud et al. 2021; Shackell et al. 2021).
In an ecosystem context, conservation and management strategies need to account for warming-induced shifts in productivity, species composition and food-web functioning, which requires comprehensive multi-species and ecosystem monitoring to inform ecosystem assessments and modeling projections (Mills et al. 2013; Bryndum-Buchholz et al. 2021). Fishers may need to diversify their catches to adapt to rapidly shifting resource composition and abundance, requiring more flexibility in licensing and quota allocation (Mills et al. 2013; Pinsky and Fogarty 2012; Pershing et al. 2021). Other possible adaptations include changes in gear use, timing and location of fishing, involvement in community co-management, or supplementing income with nonfishing activities (McClenachan et al. 2019b). An overall reduction in fishing pressure to enhance ecosystem resilience and a precautionary approach will be increasingly important to avoid unexpected sudden or cumulative consequences that may trigger species collapses or regime shifts.
Climate-smart conservation can help maintain high levels of marine biodiversity and associated ecosystem functions and services, providing a buffer against climate change impacts and an insurance for potential evolution and adaptation (Tittensor et al. 2019; Wilson et al. 2020; Lotze 2021). Both Canada and the United States (DFO 2021b; NOAA 2021b) are committed to protecting 30% of their oceans by 2030 with a mix of marine protected areas (MPAs), no-take reserves, and other area-based conservation measures. So far, the integration of climate change adaptation and resilience into marine conservation planning has been limited (Bryndum-Buchholz et al. 2022). Given current and projected climate change impacts, a proactive and climate-smart approach for marine conservation networks is advisable. This should include incorporating climate-resilience as an overall conservation objective, integrating climate change into spatial network design, and defining climate-relevant monitoring and MPA management plans (Tittensor et al. 2019; Wilson et al. 2020; Bryndum-Buchholz et al. 2022). Protected areas should include hot spots of threatened biodiversity and proper representation of different habitats, biomes, and climate change impacts. The latter should include climate refugia that are projected to experience very little change, areas with strong projected changes that would foster rapid evolution, and areas that can serve as steppingstones or corridors to support connectivity and migration. A mix of flexible and static MPAs as well as dynamic management measures and buffer zones around MPAs would also enhance adaptation potential (Tittensor et al. 2019: Wilson et al. 2020; Lotze 2021). Such climate-smart conservation can help build resilience into regional MPA networks and contribute towards global conservation goals.
To be successful, both climate-adapted management and conservation need to be backed by explicit adaptation objectives in policy and legislation providing a clear mandate, and strong governance with efficient implementation and enforcement. This requires political awareness and willingness to prepare for climate change and its consequences (Beazley and Olive 2021; Lindegren and Brander 2018; Bryndum-Buchholz et al. 2021). Currently in Canada, neither the Fisheries Act nor its recent amendment (Bill C-68 in 2019) explicitly address climate change; however, recent changes include requirements for sustainable fish stock management, rebuilding plans for depleted populations, and protection of all fish habitats, which could facilitate ecosystem-based management and climate adaptation (DFO 2019b; Bryndum-Buchholz et al. 2021). DFO has also proposed a risk-based framework to include climate change into science advice, with an initial focus on fisheries stock assessments and support of adaptive management (Duplisea et al. 2020). Recent amendments to Canada’s Oceans Act (Bill C-55 in 2019) have enhanced ecosystem and habitat protection, which may further facilitate protection and climate resilience (Bryndum-Buchholz et al. 2021). In the US with its Climate Science Strategy of 2015, NOAA Fisheries officially considers the effects of climate change on marine resources and associated management regimes and aims to enhance the production, delivery, and use of climate-related information (Link et al. 2015). The strategy’s implementation is managed through Regional Action Plans that aim to build regional capacity to address the outlined objectives (NMFS 2021). Together, climate-adapted policy, conservation, and management could create a win–win situation to sustain and enhance biodiversity protection, viable fisheries, and associated ocean benefits (Sala et al. 2021).
Caveats and next steps
One major concern in our synthesis is the many gaps in record coverage that could potentially be closed with targeted future research. Most results covering the deeper past are based on few locations (Fig. 1); additional sediment cores, microfossil sites, coral and mollusk archives, and shell midden analyses would enhance representation and resolution of different influences over space and time. A more complete reconstruction of both inshore (mollusk archives, shell middens) and offshore (coral archives, microfossils) ecosystem dynamics would also help to gain broader insight and better separate the climate and exploitation footprint. Long, sustained time series of vertically resolved temperature on the shelf would ensure that we do not have to infer bottom temperature based on surface temperature alone. Long-term records on other environmental conditions, particularly nutrients and primary production, would also be major milestones towards greater understanding of past ecosystem dynamics.
Another caveat concerns various biases in environmental and ecological archives. Microfossils and archaeological remains only contain species with hard shells, bones, teeth, scales or other structural parts that are preserved over time, and smaller, younger individuals do not preserve as well as older ones (Lotze et al. 2011). A human selection bias in archaeological and fisheries data means only species that people valued, collected, or harvested are recorded (Lotze et al. 2011). Another bias is the imprecision of radiocarbon dating, which is required to precisely connect records to past events.
Matching records from various disciplines with different units, precision, biases, and absolute and relative values present numerous challenges. We aimed to stay as true to the original data as possible and focus on general trajectories of change rather than connecting or blending all individual data sets. More research into how to best match and standardize the various records is required to derive consistent data sets and timelines, which would help to understand the overall magnitude of change. Temporally and spatially resolved timelines of human impacts, including population density, fishing effort, habitat alterations, and nutrient loading would help inform model-based resource population and ecosystem hind- and forecasting with both climate and other drivers considered. Currently, many future model projections only reflect climate change because spatio-temporal scenarios of fishing pressure or other human impacts are not yet available, although human impact scenarios are being developed (Maury et al. 2017) to be included into ensemble modeling studies (e.g., Fish-MIP).
Another caveat surrounds the use of global model projections to assess regional climate change impacts. Generally, projections based on coarse global models are less certain for coastal regions, where regional models or regional downscaling of global models that incorporate higher resolution climate and ecosystem features would be preferred (Holt et al. 2017; Laurent et al. 2021). Another caveat concerns the climate change scenarios used, which are regularly updated and improved. We used available projections based on CMIP5, yet new results based on CMIP6 with improved representation of biochemical parameters show stronger future climate and ecosystem impacts (Laurent et al. 2021; Tittensor et al. 2021). Lastly, each model has its own biases, and model projections have multiple levels of uncertainty, including scenario uncertainty, structural uncertainty, and internal variability and parameter uncertainty (Payne et al. 2016, Cheung et al. 2016). These can be partly overcome by using ensemble modeling (Tittensor et al. 2018) as done in our SSTA and Fish-MIP results above.
The next step in this line of research is to build an integrative regional end-to-end model that links a regional climate and oceanographic model with a regional biochemical model and a regional ecosystem model to capture ecosystem structure and processes more accurately at relevant management scales (Fulton et al. 2018). Ideally, this would consist of a model ensemble to include a range of possible climate and ecosystem representations and derive estimates of uncertainty. Once quantitative spatio-temporal past-to-future scenarios of climate change, fishing or other human impacts are available, this model ensemble could be used to hindcast long-term historical changes due to climate and fishing, assess individual and cumulative effects of various impacts on individual species or overall ecosystem dynamics, and project future changes under plausible climate and human impacts scenarios.
Conclusions
Our synthesis revealed a general trend of slow, late Holocene cooling of >2.5 °C over the past ∼4000 years; relatively stable climate conditions but increasing exploitation-driven changes since European arrival; and rapid, recent warming starting in the 1960s. Future projections suggest a rapid 4.5–4.8 °C warming by the end of the 21st century under a high-emissions scenario (RCP8.5), with a time of emergence from natural climate variability in 2028 and 2034 on the Scotian Shelf and Gulf of Maine, respectively. This means the region is poised to enter climate and oceanographic conditions not seen in the region over at least four millennia. Rapid warming is already accompanied by declines in primary and secondary production, shifts from subarctic to warm-water plankton communities, decreases in many cold-water species that have sustained traditional fisheries and food webs, and an influx of warm-water species from the south that are reorganizing marine ecosystems. Future ecosystem changes will strongly depend on the development of fisheries, aquaculture, and other human activities, as well as the effectiveness of management and conservation strategies. Improved climate-adaptive conservation planning and management can help mitigate some climate change impacts and support a win–win situation for marine biodiversity, ecosystem production, and service provision into the future. This requires effective climate-adapted policies and ocean governance on local, regional, and global scales.
Acknowledgements
This project has been generously sponsored by a Strategic Projects grant of the Natural Sciences and Engineering Research Council of Canada (NSERC). Additional funding is acknowledged from the Ocean Frontier Institute (Modules F and G), NSERC (by HKL, MK, EO, and KF) and the Canada Foundation for Innovation (by MBu). We thank Acadia First Nation, and especially Chief and Council, for their support of the E’se’get Project.
References
Alexander KE, Leavenworth WB, Cournane J, Cooper AB, Claesson SB, et al. 2009. Gulf of Maine cod in 1861: historical analysis of fishery logbooks, with ecosystem implications. Fish and Fisheries, 10: 428–449.
Allyn AJ, Alexander MA, Franklin BS, Massiot-Granier F, Pershing AJ, Scott JD, and Mills KE. 2020. Comparing and synthesizing quantitative distribution models and qualitative vulnerability assessments to project marine species distributions under climate change. PLoS ONE 15(4): e0231595.
Ambrose Jr. WG, Locke WL, Fisher JL, Hamilton ND, and Levitt J. 2016. Harvest of the soft-shell clam (Mya arenaria) by Malaga Island, Maine, residents from 1865 to 1912 occurred primarily in the fall and winter based on incremental growth assessment. Journal of Island & Coastal Archaeology, 11: 50–67.
Anderson SC, Lotze HK, and Shackell NL. 2008. Evaluating the knowledge base for expanding low-trophic level fisheries in Atlantic Canada. Canadian Journal for Fisheries and Aquatic Sciences, 65: 2553–2571.
Auber A, Travers-Trolet M, Villanueva MC, and Ernande B. 2015. Regime shift in an exploited fish community related to natural climate oscillations. PLoS ONE, 10(7): e0129883.
Barnhardt WA, Gehrels WR, Belknap DF, and Kelley JR. 1995. Late quaternary relative sea-level change in the Western Gulf of Maine: Evidence for a migrating glacial forebulge. Geology, 23: 317–320.
Baumgartner T. 1992. Reconstruction of the history of Pacific sardine and Northern anchovy over the past two millennia from sediments of the Santa Barbara Basin, California. California Cooperative Oceanic Fisheries Investigations Report, 33: 24–40.
Beardsley RC, and Boicourt WC. 1981. On estuarine and continental-shelf circulation in the Middle Atlantic Bight. In Evolution of Physical Oceanography. Edited by BA Warren and C Wunsch. The MIT Press, Cambridge, Massachusetts. pp. 198–233.
Beazley KF, and Olive A. 2021. Transforming conservation in Canada: shifting policies and paradigms. FACETS, 6: 1714–1727.
Bernier RY, Jamieson RE, and Moore AM. 2018. State of the Atlantic Ocean Synthesis Report. Canadian Technical Report of Fisheries and Aquatic Sciences, 3167: iii + 149 p.
Betts M. 2019. Place-making in the pretty harbour: The archaeology of Port Joli, Nova Scotia. Canadian Museum of History and University of Ottawa Press.
Betts MW, Burchell M, and Schöne BR. 2017. An Economic History of the Maritime Woodland Period in Port Joli Harbour, Nova Scotia. Journal of the North Atlantic, 10(10): 18–42.
Betts MW, and Hrynick MG. 2021. The Archaeology of the Atlantic Northeast. University of Toronto Press, Toronto.
Betts MW, Maschner HDG, and Clark DS. 2011. Zooarchaeology of the “fish that stops” using archaeofaunas to construct long-term time series of Atlantic and Pacific cod populations. In The Archaeology of North Pacific Fisheries. Edited by M Moss and A Cannon. University of Alaska Press, Anchorage. pp. 171–195.
Betts MW, Noël S, Tourigny E, Burns M, Pope PE, and Cumbaa SL. 2014. Zooarchaeology of the historic cod fishery in Newfoundland and Labrador, Canada. Journal of the North Atlantic, 24: 1–21.
Bianucci L, Fennel K, Chabot D, Shackell N, and Lavoie D. 2015. Ocean biogeochemical models as management tools: a case study for Atlantic wolffish and declining oxygen. ICES Journal of Marine Science, 73(2): 263–274.
Bograd SJ, Checkley DA, and Wooster WS. 2003. CalCOFI: A half century of physical, chemical, and biological research in the California Current System. Deep Sea Research Part II: Topical Studies in Oceanography, 50(14–16): 2349–2353.
Bourque BJ, Johnson B, and Steneck RS. 2008. Possible prehistoric fishing effects on coastal marine food webs in the Gulf of Maine. In Human Impacts on Ancient Marine Ecosystems. Editors TC Rick and JM Erlandson. University of California Press, Berkley. pp. 165–185.
Boyce DG, Lewis MR, and Worm B. 2010. Global phytoplankton decline over the past century. Nature, 466: 591–596.
Brander KM. 2007. Global fish production and climate change. Proceedings of the National Academy of Sciences, 104(50): 19709–19714.
Brander KM. 2018. Climate change not to blame for cod population decline. Nature Sustainability, 1:262–264.
Brassell SC, Eglinton G, Marlowe IT, Pflaumann U, and Sarnthein M. 1986. Molecular stratigraphy: a new tool for climatic assessment. Nature, 320(6058): 129–133.
Brickman D, Alexander MA, Pershing A, Scott JD, and Wang Z 2021. Projections of physical conditions in the Gulf of Maine in 2050. Elementa Science of the Anthropocene, 9: 1.
Brickman D, Hebert D, and Wang Z. 2018. Mechanism for the recent ocean warming events on the Scotian shelf of eastern Canada. Continental Shelf Research, 156: 11–22.
Britten GL, Dowd M, and Worm B. 2016. Changing recruitment capacity in global fish stocks. Proceedings of the National Academy of Sciences, 113(1): 134–139.
Bryndum-Buchholz A, Boerder H, Stanley RRE, Hurley I, Boyce DG, Dunmall KM, et al. 2022. A climate-resilient marine conservation network for Canada. Facets, 7: 571–590.
Bryndum-Buchholz A, Boyce DG, Tittensor DP, Christensen V, Bianchi D, and Lotze HK. 2020. Climate change impacts and fisheries management challenges in the North Atlantic Ocean. Marine Ecology Progress Series, 648: 1–17.
Bryndum-Buchholz A, Tittensor DP, and Lotze HK. 2021. The status of climate change adaptation in fisheries management: policy, legislation, and implementation. Fish and Fisheries, 22(6): 1248–1273.
Bundy A, Gomez C, and Cook AM. 2017. Guidance framework for the selection and evaluation of ecological indicators. Canadian Technical Report of Fisheries and Aquatic Sciences, 3232: xii + 212 p.
Burchell M, Betts MW, Patton AK, and Schöne BR. 2014. Preliminary analysis of stable oxygen isotopes and shell growth in the soft-shelled clam Mya arenaria: Implications for interpreting seasonality and shellfish harvesting in Port Joli, Nova Scotia. North Atlantic Archaeology, 3: 93–108.
Busch DS, Griffis R, Link J, Abrams K, Baker J, Brainard RE, et al. 2016. Climate science strategy of the US National Marine Fisheries Service. Marine Policy, 74: 58–67.
Caesar L, McCarthy GD, Thornalley DJR, Cahill N, and Rahmstorf S. 2021. Current Atlantic meridional overturning circulation weakest in last millennium. Nature Geoscience, 14(3): 118–120.
Caesar L, Rahmstorf S, Robinson A, Feulner G, and Saba V. 2018. Observed fingerprint of a weakening Atlantic Ocean overturning circulation. Nature, 556(7700): 191–196.
Carson S, Shackell N, and Mills Flemming J. 2017. Local overfishing may be avoided by examining parameters of a spatio-temporal model. PLoS ONE, 12(9): e0184427.
Casault B, Johnson C, Devred E, Head E, Cogswell A, and Spry J. 2020. Optical, chemical, and biological oceanographic conditions on the Scotian Shelf and in the eastern Gulf of Maine during 2019. DFO Canadian Science Advisory Secretariat Research Document, 2020/071: v + 64 p.
Cheung WWL, Frölicher TL, Asch RG, Jones MC, Pinsky ML, Reygondeau G, et al. 2016. Building confidence in projections of the responses of living marine resources to climate change. ICES Journal of Marine Science, 73: 1283–1296.
Christensen, V, Guenette S, Heymans JJ, Walters CJ, Watson R, Zeller D, and Pauly D. 2003. Hundred-year decline of North Atlantic predatory fishes. Fish and Fisheries, 4: 1–24.
Claret M, Galbraith ED, Palter JB, Bianchi D, Fennel K, Gilbert D, and Dunne JP. 2018. Rapid coastal deoxygenation due to ocean circulation shift in the northwest Atlantic. Nature Climate Change, 8(10): 868–872.
Cramer K, O’Dea A, Clark TR, Zhao J, and Norris RD. 2017. Prehistorical and historical declines in Caribbean coral reef accretion rates driven by loss of parrotfish. Nature Communications, 8: 14160.
Deser C, Alexander MA, Xie S-P, and Phillips AS. 2010. Sea surface temperature variability: Patterns and mechanisms. Annual Review of Marine Science, 2: 115–143.
Dever M, Hebert D, Greenan BJ, Sheng J, and Smith PC. 2016. Hydrography and coastal circulation along the Halifax Line and the connections with the Gulf of St Lawrence. Atmosphere-Ocean, 54(3): 199–217.
DFO. 2019a. Framework for Incorporating Climate-Change Considerations into Fisheries Stock Assessments. Department of Fisheries and Oceans Canada, Retrieved from dfo-mpo.gc.ca/csas-sccs/Publications/SAR-AS/2019/2019_029-eng.Pdf.
DFO. 2019b. Fisheries Act updates to better protect fish and fish habitat. [online]: Available from dfo-mpo.gc.ca/campaign-campagne/fisheries-act-loi-sur-les-peches/intro ducti on-eng.html.
DFO. 2021a. Marine environmental data section archive. Ecosystem and Oceans Science, Department of Fisheries and Oceans Canada. [online]: Available from meds-sdmm.dfo-mpo.gc.ca.
DFO. 2021b. Reaching Canada’s marine conservation targets. [online]: Available from dfo-mpo.gc.ca/oceans/conservation/plan/index-eng.html.
Drinkwater KF, Petrie B. 2011. A note on the long-term sea surface temperature records at Boothbay Harbor, Maine. Journal of Northwest Atlantic Fisheries Science, 43: 93–101.
Dunn DC, Maxwell SM, Boustany AM, and Halpin PN. 2016. Dynamic ocean management increases the efficiency and efficacy of fisheries management. Proceedings of the National Academy of Sciences, 113(3): 668–673.
Duplisea DE, Roux M-J, Hunter KL, and Rice J. 2020. Resource management under climate change: a risk-based strategy to develop climate-informed science advice. DFO Canadian Science Advisory Secretariat Research Document, 2019/044.
Dusseault M. 2021. Investigating variability in marine mollusk carbonates using infrared spectroscopy and radiocarbon analysis. M.Sc. thesis, Memorial University of Newfoundland, St. John’s, Newfoundland, Canada.
Edgecombe RB, Scott DB, and Fader GBJ. 1999. New data from Halifax Harbour: paleoenvironment and a new Holocene sea-level curve for the inner Scotian Shelf. Canadian Journal of Earth Sciences, 36: 805–817.
Etayo-Cadavid M, Andrus C, Jones K, and Hodgins G. 2019. Subseasonal variations in marine reservoir age from pre-bomb Donax obesulus and Protothaca asperrima shell carbonate. Chemical Geology, 526: 110–116.
Fennel K. 2010. The role of continental shelves in nitrogen and carbon cycling: Northwestern North Atlantic case study. Ocean Science, 6: 539–548.
Ferguson J, Johnson K, Santos G, Meyer L, and Tripati A. 2013. Investigating δ13C and Δ14C within Mytilus californianus shells as proxies of upwelling intensity. Geochemistry, Geophysics, Geosystems, 14(6): 1856–1865.
Filippova A, Kienast M, Frank M, and Schneider RR. 2016. Alkenone paleothermometry in the North Atlantic: A review and synthesis of surface sediment data and calibrations. Geochemistry, Geophysics, Geosystems, 17(4): 1370–1382.
Finney BP, Alheit J, Emeis K-C, Field DB, Gutiérrez D, and Struck U. 2010. Paleoecological studies on variability in marine fish populations: a long-term perspective on the impacts of climatic change on marine ecosystems. Journal of Marine Systems, 79(3-4): 316–326.
Frank KT, Petrie B, Choi JS, and Leggett WC. 2005. Trophic cascades in a formerly cod-dominated ecosystem. Science, 308(5728): 1621–1623.
Frank KT, Petrie B, Fisher JAD, and Leggett WC. 2011. Transient dynamics of an altered large marine ecosystem. Nature, 477(7362): 86–89.
Free CM, Thorson JT, Pinsky ML, Oken KL, Wiedemann J, and Jensen OP. 2019. Impacts of historical warming on marine fisheries production. Science, 363(6430): 979–983.
Freeman E, Woodruff SD, Worley SJ, Lubker SJ, Kent EC, Angel WE, et al. 2017. ICOADS Release 3.0: a major update to the historical marine climate record. International Journal of Climatology, 37(5): 2211–2232.
Friedland KD, Langan JA, Large SI, Selden RL, Link JS, Watson RA, and Collie JS. 2019. Changes in higher trophic level productivity, diversity and niche space in a rapidly warming continental shelf ecosystem. Science of The Total Environment, 704: 135270.
Friedland KD, Morse RE, Shackell N, Tam JC, Morano JL, Moisan JR, and Brady DC. 2020. Changing physical conditions and lower and upper trophic level responses on the US Northeast Shelf. Frontiers in Marine Science, 7: 567445.
Fulton EA, Hobday AJ, Pethybridge H, Blanchard J, Bulman C, Butler I, et al. 2018. Decadal scale projection of changes in Australian fisheries stocks under climate change. CSIRO Report to FRDC Project No. 2016/139, Fisheries Research and Development Corporation, Hobart, TAS, Australia.
Galbraith ED, Carozza DA, and Bianchi D. 2017. A coupled human-Earth model perspective on long-term trends in the global marine fishery. Nature Communication, 8: 14884.
Giese BS, Seidel HF, Compo GP, and Sardeshmukh PD. 2016. An ensemble of ocean reanalyses for 1815-2013 with sparse observational input. Journal of Geophysical Research: Oceans, 121(9): 6891–6910.
Greenan BJW, Shackell NL, Ferguson K, Greyson P, Cogswell A, Brickman D, et al. 2019. Climate change vulnerability of American lobster fishing communities in Atlantic Canada. Frontiers in Marine Science, 6: 579.
Greene CH, and Pershing AJ. 2000. The response of Calanus finmarchicus populations to climate variability in the Northwest Atlantic: basin-scale forcing associated with the North Atlantic Oscillation. ICES Journal of Marine Science, 57(6): 1536–1544.
Greene CH, and Pershing AJ. 2003. The flip-side of the North Atlantic Oscillation and modal shifts in slope-water circulation patterns. Limnology and Oceanography, 48(1): 319–322.
Grieve BD, Hare JA, and Saba VS. 2017. Projecting the effects of climate change on Calanus finmarchicus distribution within the US Northeast Continental Shelf. Scientific Reports 7(1): 1–12.
Grossman EL, and Ku T-L. 1986. Oxygen and carbon isotope fractionation in biogenic aragonite: temperature wffects. Chemical Geology, 59: 59–74.
Han G, and Loder JW. 2003. Three-dimensional seasonal-mean circulation and hydrography on the eastern Scotian Shelf. Journal of Geophysical Research: Oceans, 108(C5): 1–21.
Harrison WG, and Li WKW. 2008. Phytoplankton growth and regulation in the Labrador Sea: light and nutrient limitation. Journal of Northwest Atlantic Fisheries Science, 39: 71–82.
Hátún H, Azetsu-Scott K, Somavilla R, Rey F, Johnson C, Mathis M, et al. 2017. The subpolar gyre regulates silicate concentrations in the North Atlantic. Scientific Reports, 7: 14576.
Heaton T, Köhler P, Butzin M, Bard E, Reimer R, Austin W, et al. 2020. Marine 20-The marine radiocarbon age calibration curve (0-55,000 cal BP). Radiocarbon, 62(4): 779–820.
Hebert D, Layton C, Brickman D, and Galbraith PS. 2021. Physical Oceanographic Conditions on the Scotian Shelf and in the Gulf of Maine during 2019. DFO Canadian Science Advisory Secretariat Research Document, 2021/040. iv + 58 p.
Henson SA, Beaulieu C, Ilyina T, John JG, Long M, Séférian R, et al. 2017. Rapid emergence of climate change in environmental drivers of marine ecosystems. Nature Communications, 8: 14682.
Hirahara S, Ishii M, and Fukuda Y. 2014. Centennial-scale sea surface temperature analysis and its uncertainty. Journal of Climate, 27(1): 57–75.
Holt J, Hyder P, Ashworth M, Harle J, Hewitt HT, Liu H, et al. 2017. Prospects for improving the representation of coastal and shelf seas in global ocean models. Geoscientific Model Development, 10: 499–523.
Huang B, Thorne PW, Banzon VF, Boyer T, Chepurin G, Lawrimore JH, et al. 2017. Extended reconstructed sea surface temperature, version 5 (ERSSTv5): upgrades, validations, and intercomparisons. Journal of Climate, 30(20): 8179–8205.
Jackson JBC, Kirby MX, Berger WH, Bjorndal KA, Botsford LW, Bourque BJ, et al. 2001. Historical overfishing and the recent collapse of coastal ecosystems. Science, 293(5530): 629–637.
Jenkins WJ, Smethie WM, Boyle EA, and Cutter GA 2015. Water mass analysis for the U.S. GEOTRACES (GA03) North Atlantic sections. Deep-Sea Research Part II, 116: 6–20.
Jones EP, Swift JH, Anderson LG, Lipizer M, Civitarese G, Falkner KK, et al. 2003. Tracing Pacific water in the North Atlantic Ocean. Journal of Geophysical Research, 108: 3116.
Jones MC, and Cheung WWL. 2015. Multi-model ensemble projections of climate change effects on global marine biodiversity. ICES Journal of Marine Science, 72(3): 741–752.
Jones WA, and Checkley DM. 2019. Mesopelagic fishes dominate otolith record of past two millennia in the Santa Barbara Basin. Nature Communications, 10: 4564.
Kay JE, Deser C, Phillips A, Mai A, Hannay C, Strand G, et al. 2015. The Community Earth System Model (CESM) large ensemble project: A community resource for studying climate change in the presence of internal climate variability. Bulletin of the American Meteorological Society, 96(8): 1333–1349.
Keigwin LD, and Jones GA. 1995. The marine record of deglaciation from the continental margin off Nova Scotia. Paleoceanography, 10: 973–985.
Keigwin LD, and Pilskaln CH. 2015. Sediment flux and recent paleoclimate in Jordan Basin, Gulf of Maine. Continental Shelf Research, 96: 45–55.
Keigwin LD, Sachs JP, and Rosenthal Y. 2003. A 1600-year history of the Labrador Current off Nova Scotia. Climate Dynamics, 21: 53–62.
Keigwin LD, Sachs JP, Rosenthal Y, and Boyle EA. 2005. The 8200 year B.P. event in the slope water system, western subpolar North Atlantic. Paleoceanography, 20: PA2003.
Kelley JT, Belknap DF, Kelley AR, and Claesson SH. 2013. A model for drowned terrestrial habitats with associated archeological remains in the Northwestern Gulf of Maine. Marine Geology, 338: 1–16.
Kenchington TJ, and Kenchington EL. 1993. An eighteenth century commercial length-frequency sample of Atlantic Cod, Gadus morhua, based on archaeological data. Fisheries Research, 18: 335–347.
Kleisner KM, Fogarty MJ, McGee S, Hare JA, Moret S, Perretti CT, and Saba VS. 2017. Marine species distribution shifts on the US Northeast Continental Shelf under continued ocean warming. Progress in Oceanography 153: 24–36.
Kwiatkowski L, Aumont O, and Bopp L. 2019. Consistent trophic amplification of marine biomass declines under climate change. Global Change Biology, 25(1): 218–229.
Laloyaux P, de Boisseson E, Balmaseda M, Bidlot J-R, Broennimann S, Buizza R, et al. 2018. CERA-20C: A coupled reanalysis of the twentieth century. Journal of Advances in Modeling Earth Systems, 10(5): 1172–1195.
Laurent A, Fennel K, and Kuhn A. 2021. An observation-based evaluation and ranking of historical Earth System Model simulations for regional downscaling in the northwest North Atlantic Ocean. Biogeosciences, 18: 1803–1822.
Lavoie D, Lambert N, and Gilbert D. 2017. Projections of future trends in biogeochemical conditions in the northwest Atlantic using CMIP5 Earth system models. Atmosphere-Ocean, 57(1): 18–40.
Lear WH. 1998. History of fisheries in the Northwest Atlantic: the 500-year perspective. Journal of Northwest Atlantic Fishery Science, 23: 41–74.
Leduc G, Schneider RR, Kim J-H, and Lohmann G. 2010. Holocene and Eemian Sea surface temperature trends as revealed by alkenone and Mg/Ca paleothermometry. Quaternary Science Reviews, 29(7-8): 989–1004.
Levac E. 2001. High resolution Holocene palynological record from the Scotian Shelf. Marine Micropaleontology, 43(3–4): 179–197.
Lindegren M, and Brander K. 2018. Adapting fisheries and their management to climate change: a review of concepts, tools, frameworks, and current progress toward implementation. Reviews in Fisheries Science & Aquaculture, 26(3): 400–415.
Lindegren M, Diekmann R, and Möllmann C. 2010. Regime shifts, resilience and recovery of a cod stock. Marine Ecology Progress Series 402: 239–253.
Link JS, Griffis R, and Busch S. 2015. NOAA Fisheries Climate Science Strategy, NMFS-F/SPO-155. National Marine Fisheries Service. [online]: Available from st.nmfs.noaa.gov/Assets/ecosystems/climate/documents/NCSS_Final.pdf.
Loder JW and Petrie B. 1998. The Coastal Ocean off Northeastern North America A large-scaleview. This sea, 11: 105–133.
Loder JW, van der Baaren A, and Yashayaev I. 2015. Climate comparisons and change projections for the Northwest Atlantic from six CMIP5 models. Atmosphere-Ocean, 53(5): 529–555.
Lotze HK. 2021. Primer marine biodiversity conservation. Current Biology, 31: R1141–R1224.
Lotze HK, and McClenachan L. 2014. Marine Historical Ecology: informing the future by learning from the past. In Marine Community Ecology and Conservation. Edited by MB Bertness, BR Silliman, JF Bruno, and JJ Stachowicz. Sinauer. pp. 165–200.
Lotze HK, and Milewski I. 2002. Two hundred years of ecosystem and food web changes in the Quoddy region, outer Bay of Fundy. Conservation Council of New Brunswick, Fredericton, New Brunswick, Canada, 188 p.
Lotze HK, and Milewski I. 2004. Two centuries of multiple human impacts and successive changes in a North Atlantic food web. Ecological Applications, 14(5): 1428–1447. [online]: Available from jstor.org/stable/4493661.
Lotze HK, and Worm B. 2009. Historical baselines for large marine animals. Trends in Ecology & Evolution, 24(5): 254–262.
Lotze HK, Bryndum-Buchholz A, and Boyce DG. 2021. Effects of climate change on food production (Fishing). In The Impacts of Climate Change: Comprehensive Study of the Physical, Societal and Political Issues. 1st ed. Edited by T Letcher. Elsevier. pp. 205–231.
Lotze HK, Erlandson JM, Newman MJ, Norris RD, Roy K, Smith TD, and Whitcraft C. 2011. Uncovering the ocean’s past. In Shifting Baselines: The Past and the Future of Ocean Fisheries. Edited by JBC Jackson, E Sala, and K Alexander. Island Press. pp. 137–162.
Lotze HK, Lenihan HS, Bourque BJ, Bradbury RH, Cooke RG, Kay MC, et al. 2006. Depletion, degradation, and recovery potential of estuaries and coastal seas. Science, 312(5781): 1806–1809.
Lotze HK, Tittensor DP, Bryndum-Buchholz A, Eddy TD, Cheung WWL, Galbraith ED, et al. 2019. Global ensemble projections reveal trophic amplification of ocean biomass declines with climate change. Proceedings of the National Academy of Sciences USA, 116(26): 12907–12912.
Lotze HK. 2010. Historical reconstruction of human-induced changes in U.S. estuaries. Oceanography and Marine Biology: An Annual Review, 48: 265–336.
Marchitto TM, and deMenocal P. 2003. Late Holocene variability of upper North Atlantic deep water temperature and salinity. Geochemistry, Geophysics, Geosystems 4(12): 1100.
Maureaud AA, Frelat R, Pecuchet L, Shackell N, Merigot B, Pinsky ML, et al. 2021. Are we ready to track climate-driven shifts in marine species across international boundaries? - A global survey of scientific bottom trawl data. Global Change Biology, 27: 220–236.
Maury O, Campling L, Arrizabalaga H, Aumont O, Bopp L, Merino H, et al. 2017. From shared socio-economic pathways (SSPs) to oceanic system pathways (OSPs): building policy-relevant scenarios for global oceanic ecosystems and fisheries. Global Environmental Change, 45: 203−216.
McClenachan, L, Grabowski, JH, Marra, M, McKeon, CS, Neal, BP, Record, NR, and Scyphers, SB. 2019a. Shifting perceptions of rapid temperature changes’ effects on marine fisheries, 1945–2017. Fish and Fisheries 20(6): 1111–1123.
McClenachan, L, Scyphers, S, and Grabowski, JH. 2019b. Views from the dock: Warming waters, adaptation, and the future of Maine’s lobster fishery. Ambio 49(1): 144–155. hdl.handle.net/2047/D20325645.
Mellon S, Kienast M, Algar C, deMenocal P, Kienast SS, Marchitto T, and Thomas H. 2019. Foraminifera trace anthropogenic CO2 in the NW Atlantic by 1950. Geophysical Research Letters, 46(24): 14683–14691.
Miller AAL, Mudie PJ, and Scott DB. 1982. Holocene history of Bedford Basin, Nova Scotia: foraminifera, dinoflagellate, and pollen records. Canadian Journal of Earth Sciences, 19(12): 2342–2367.
Mills KE, Pershing AJ, Brown CJ, Chen Y, Chiang F-S, Holland DS, et al. 2013. Fisheries management in a changing climate: Lessons from the 2012 ocean heat wave in the Northwest Atlantic. Oceanography, 26(2): 191–195.
Monge Soares AM, and Alveirinho Dias JM. 2006. Coastal upwelling and radiocarbon-evidence for temporal fluctuations in ocean reservoir effect off Portugal during the Holocene. Radiocarbon, 48(1): 45–60.
Mudie PJ, Rochon A, and Levac E. 2002. Palynological records of red tide-producing species in Canada: past trends and implications for the future. Palaeogeography, Palaeoclimatology, Palaeoecology, 180(1–3): 159–186.
Myers RA, Hutchings J, and Barrowman N. 1997. Why do fish stocks collapse? The example of cod in Atlantic Canada. Ecological Applications, 7(1): 91–106.
Myers RA, and Worm B. 2005. Extinction, survival or recovery of large predatory fishes. Philosophical Transactions of the Royal Society B: Biological Sciences, 360(1453): 13–20.
NFMS. 2021. Climate Science Strategy Regional Action Plans. [online]: Available from fisheries.noaa.gov/national/climate/climate-science-strategy-regional-action-plans.
NOAA. 2019. Current Conditions of the Northeast U.S. Shelf Ecosystem: Fall 2019 Update. A summary of temperature, salinity, chlorophyll and other conditions on the Northeast U.S. Continental Shelf during the first half of 2019. National Oceanographic and Atmospheric Administration, USA. [online]: Available from fisheries.noaa.gov/new-england-mid-atlantic/ecosystems/current-conditions-northeast-us-shelf-ecosystem-fall-2019.
NOAA. 2021a. Current Conditions of the Northeast U.S. Shelf Ecosystem: Spring 2021 Update. A summary of temperature, chlorophyll and other oceanographic conditions on the Northeast U.S. Continental Shelf during the last half of 2020. National Oceanographic and Atmospheric Administration, USA. [online]: Available from fisheries.noaa.gov/new-england-mid-atlantic/climate/current-conditions-northeast-us-shelf-ecosystem-spring-2021-update.
NOAA. 2021b. Understanding Area-based Management in U.S. Waters. Context for the 30 × 30 marine conservation goal. National Marine Protected Areas Center, NOAA, USA. [online]: Available from marineprotectedareas.noaa.gov/gallery/understanding-area-based-mgmt-in-us-waters.htm
Pauly D. 1995. Anecdotes and the shifting baseline syndrome of fisheries. Trends in Ecology & Evolution, 10(10): 430.
Payne MR, Barange M, Cheung WWL, MacKenzie BR, Batchelder HP, Cormon X, et al. 2016. Uncertainties in projecting climate change impacts in marine ecosystems. ICES Journal of Marine Science, 73: 1272–1282.
Pedersen EJ, Thompson PL, Ball RA, Fortin M-J, Gouhier TC, Link H, et al. 2017. Signatures of the collapse and incipient recovery of an overexploited marine ecosystem. Royal Society Open Science, 4: 170215.
Pedersen JST, van Vuuren DP, Aparício BA, Swart R, Gupta J, and Santos FD. 2020. Variability in historical emissions trends suggests a need for a wide range of global scenarios and regional analyses. Communications Earth & Environment 1: 41.
Pepin P, King J, Holt C, Gurney-Smith H, Shackell N, Hedges K, and Bundy A. 2020. Incorporating climate, oceanographic and ecological change considerations into population assessments: A review of Fisheries and Oceans Canada’s science advisory process. DFO Canadian Science Advisory Secretariat Research Document, 2019/043.
Pershing AJ, Alexander MA, Brady DC, Brickman D, Curchitser EN, Diamond AW, et al. 2021. Climate impacts on the Gulf of Maine ecosystem: A review of observed and expected changes in 2050 from rising temperatures. Elementa Science of the Anthropocene, 9: 1.
Pershing AJ, Alexander MA, Hernandez CM, Kerr LA, Le Bris A, Mills KE, et al. 2015. Slow adaptation in the face of rapid warming leads to collapse of the Gulf of Maine cod fishery. Science, 350(6262): 809–812.
Petrie B, and Drinkwater K. 1993. Temperature and salinity variability on the Scotian Shelf and in the Gulf of Maine 1945-1990. Journal of Geophysical Research: Oceans, 98: 79–89.
Petrie B, and Yeats P. 2000. Annual and interannual variability of nutrients and their estimated fluxes in the Scotian Shelf – Gulf of Maine region. Canadian Journal of Fisheries and Aquatic Sciences, 57: 2536–2546.
Pinsky ML, and Fogarty M. 2012. Lagged social-ecological responses to climate and range shifts in fisheries. Climate Change 115: 883–891.
Pinsky ML, Worm B, Fogarty MJ, Sarmiento JL, and Levin SA. 2013. Marine taxa track local climate velocities. Science 341: 1239–12342.
Prahl FG, Muehlhausen LA, and Zahnle DL. 1988. Further evaluation of long-chain alkenones as indicators of paleoceanographic conditions. Geochimica et Cosmochimica Acta, 52(9): 2303–2310.
Rayner NAA, Parker DE, Horton EB, Folland CK, Alexander LV, Rowell DP, et al. 2003. Global analyses of sea surface temperature, sea ice, and night marine air temperature since the late nineteenth century. Journal of Geophysical Research: Atmospheres, 108(D14): 4407.
Reygondeau G, and Beaugrand G. 2011. Future climate driven shifts in distribution of Calanus finmarchicus. Global Change Biology, 17: 756–766.
Riahi K, Rao S, Krey V, Cho C, Chirkov V, Fischer G, et al. 2011. RCP 8.5—A scenario of comparatively high greenhouse gas emissions. Climatic Change, 109(1–2): 33.
Robinson LF, Adkins JF, Frank N, Gagnon AC, Prouty NG, Roark EB, and Van de Flierdt T. 2014. The geochemistry of deep-sea coral skeletons: a review of vital effects and applications for palaeoceanography. Deep Sea Research Part II, 99: 184–198.
Rojo A. 1986. Live length and weight of cod (Gadus morhua) estimated from various skeletal elements. North American Archaeologist, 7: 329–351.
Rojo A. 1987. Excavated fish vertebrae as predictors in bioarchaeological research. North American Archaeologist, 8: 209–226.
Rojo A. 1990. Faunal analysis of fish remains from Cellar’s Cove, Nova Scotia. Archaeology of Eastern North America, 18: 89–108.
Rose GA. 2004. Reconciling overfishing and climate change with stock dynamics of Atlantic cod (Gadus morhua) over 500 years. Canadian Journal of Fisheries and Aquatic Sciences, 61(9): 1553–1557.
Rosenberg AA, Bolster WJ, Alexander KE, Leavenworth WB, Cooper AB, and McKenzie MG. 2005. The history of ocean resources: modelling cod biomass using historical records. Frontiers in Ecology and the Environment, 3(2): 84–90.
Ross CH, Record NR, Pendleton DE, Tupper B, and Brickman D. 2021. Predicting regions of North Atlantic right whale, Eubalaena glacialis, habitat suitability in the Gulf of Maine in 2050. Elementa: Science of the Anthropocene 9(1): 00058.
Rutherford K. 2021. Source or sink? A modeling study of inorganic carbon cycling on the Scotian Shelf. Doctoral Thesis, Dalhousie University, Halifax, Nova Scotia, Canada. 160 p.
Rutherford K, and Fennel K. 2018. Diagnosing transit times on the northwestern North Atlantic continental shelf. Ocean Science, 14(5): 1207–1221.
Saba VS, Griffies SM, Anderson WG, Winton M, Alexander MA, Delworth TL, et al. 2016. Enhanced warming of the Northwest Atlantic Ocean under climate change. Journal of Geophysical Research: Oceans, 121(1): 118–132.
Sachs JP. 2007. Cooling of Northwest Atlantic slope waters during the Holocene. Geophysical Research Letters, 34: L03609.
Sala E, Mayorga J, Bradley D, Cabral RB, Atwood TB, Auber A, et al. 2021. Protecting the global ocean for biodiversity, food and climate. Nature, 592: 397–402.
Salisbury JE, Jönsson BF. 2018. Rapid warming and salinity changes in the Gulf of Maine alter surface ocean carbonate parameters and hide ocean acidification. Biogeochemistry, 141: 401–418.
Schnitker D, and Jorgensen JB. 1990. Late Glacial and Holocene Diatom Successions in the Gulf of Maine: Response to Climatologic and Oceanographic Change. In Evolutionary Biogeography of the Marine Algae of the North Atlantic. Edited by DJ Garbary and GR South. NATO ASI Series (Series G: Ecological Sciences), Vol 22. Springer, Berlin, Heidelberg.
Scott DB, Mudie PJ, Vilks G, and Younger DC. 1984. Latest Pleistocene—Holocene paleoceanographic trends on eastern Canadian margin: Foraminiferal, dinoflagellate and pollen evidence. Marine Micropaleontology, 9(3): 181–218.
Shackell NL, Fisher JAD, Den Heyer CE, Hennen DR, Seitz AC, Le Bris A, et al. 2021. Spatial ecology of Atlantic halibut across the Northwest Atlantic: A recovering species in an era of climate change. Reviews in Fisheries Science & Aquaculture, published online: 1–25.
Shackell NL, Ricard D, and Stortini CH. 2014. Thermal habitat index of many Northwest Atlantic temperate species stays neutral under warming projected for 2030 but changes radically by 2060. PLoS ONE, 9(3): e90662.
Shan S, Sheng J, Ohashi K, Dever M. 2016. Assessing the performance of a multi-nested ocean circulation model using satellite remote sensing and in-situ observations. Satellite Oceanography and Meteorology, 1(1): 39–59.
Shaw J, Fader GB, and Taylor RB. 2009. Submerged early Holocene coastal and terrestrial landforms on the inner shelves of Atlantic Canada. Quaternary International, 206(1–2): 24–34.
Shaw J, Gareau P, and Courtney RC. 2002. Palaeogeography of Atlantic Canada 13–0 kyr. Quaternary Science Reviews, 21(16–17): 1861–1878.
Sherwood OA, Davin SH, Lehmann N, Buchwald C, Edinger EN, Lehmann MF, and Kienast M. 2021. Stable isotope ratios in seawater nitrate reflect the influence of Pacific water along the northwest Atlantic margin. Biogeosciences, 18: 4491–4510.
Sherwood OA, and Edinger EN. 2009. Ages and growth rates of some deep-sea gorgonian and antipatharian corals of Newfoundland and Labrador. Canadian Journal of Fisheries and Aquatic Sciences, 66(1): 142–152.
Sherwood OA, Lehmann MF, Schubert CJ, Scott DB, and McCarthy MD. 2011. Nutrient regime shift in the western North Atlantic indicated by compound-specific δ15N of deep-sea gorgonian corals. Proceedings of the National Academy of Sciences of the United States of America, 108(3): 1011–1015.
Sherwood OA, Scott DB, and Risk MJ. 2006. Late Holocene radiocarbon and aspartic acid racemization dating of deep-sea octocorals. Geochimica et Cosmochimica Acta, 70(11): 2806–2814.
Sibert EC, Cramer KL, Hastings PA, and Norris RD. 2017. Methods for isolation and quantification of microfossil fish teeth and elasmobranch dermal denticles (ichthyoliths) from marine sediments. Palaeontologia Electronica 20(1): 1–14.
Spiess AE, Trautman E, and Kupferschmid T. 1990. Prehistoric occupation at Reversing Falls. Unpublished file report, Maine Historic Preservation Commission, Augusta, Maine. 1–28.
Steneck RS. 1997. Fisheries-induced biological changes to the structure and function of the Gulf of Maine ecosystem. Regional Association for the Research on the Gulf of Maine (RARGOM) report 91-1, Hanover, New Hampshire, USA.
Stortini CH, Shackell NL, Tyedmers P, and Beazley K. 2015. Assessing marine species vulnerability to projected warming on the Scotian Shelf, Canada. ICES Journal of Marine Science, 72(6): 1731–1743.
Thibodeau B, Not C, Hu J, Schmittner A, Noone D, Tabor C, et al. 2018. Last century warming over the Canadian Atlantic shelves linked to weak Atlantic Meridional Overturning Circulation. Geophysical Research Letters, 45(22): 12,376–12,385.
Thornalley DJR, Oppo DW, Ortega P, Robson JI, Brierley CM, Davis R, et al. 2018. Anomalously weak Labrador Sea convection and Atlantic overturning during the past 150 years. Nature, 556(7700): 227–230.
Tittensor DP, Beger M, Börder K, Boyce D, Cavanagh R, Cosandey-Godin A, et al. 2019. Integrating climate adaptation and biodiversity conservation in the global ocean. Science Advances, 5(11): eaay9969. [online]: Available from advances.sciencemag.org/content/5/11/eaay9969.
Tittensor DP, Eddy TD, Lotze HK, Galbraith ED, Cheung WWL, Barange M, et al. 2018. A protocol for the intercomparison of marine fishery and ecosystem models: Fish-MIP v1.0. Geoscientific Model Development, 11: 1421–1442.
Tittensor DP, Novaglio C, Harrison CS, Heneghan RF, Barrier N, Bianchi D, et al. 2021. Next-generation Earth system models reveal higher climate risks for marine ecosystems. Nature Climate Change, 11: 973–981.
Townsend DW, Pettigrew NR, Thomas MA, Neary MG, McGillicuddy J, Dennis J, and O’Donnell J. 2015. Water masses and nutrient sources to the Gulf of Maine. Journal of Marine Research, 73(3–4): 93–122.
Townsend DW, Rebuck ND, Thomas MA, Karp-Boss L, and Gettings RM. 2010. A changing nutrient regime in the Gulf of Maine. Continental Shelf Research, 30(7): 820–832.
Tremblay J-É, Anderson LG, Matrai P, Couple P, Bélanger S, Michel C, and Reigstad M. 2015. Global and regional drivers of nutrient supply, primary production and CO2 drawdown in the changing Arctic Ocean. Progress in Oceanography, 139: 171–196.
Vacchi M, Engelhart SE, Nikitina D, Ashe EL, Peltier WR, Roy K, et al. 2018. Postglacial relative sea-level histories along the eastern Canadian coastline. Quaternary Science Reviews, 201: 124–146.
VanderZwaag, DL, Bailey, M, and Shackell, NL. 2017. Canada–US fisheries management in the Gulf of Maine: Taking stock and charting future coordinates in the face of climate change. Ocean Yearbook Online 31: 1–26.
Waller JD, Wahle RA, McVeigh H, and Fields DM. 2017. Linking rising pCO2 and temperature to the larval development and physiology of the American lobster (Homarus americanus). ICES Journal of Marine Science, 74:1210–1219.
Wanamaker AD Jr., Kreutz KJ, Borns HW, Introne DS, Feindel S, Funder S, et al. 2007. Experimental Determination of Salinity, Temperature, Growth, and Metabolic Effects on the Shell Isotope Chemistry of Mytilus edulis Collected from Maine and Greenland. Paleoceanography, 22:1–12.
Wanamaker Jr. AD, Kreutz KJ, Schöne BR, and Introne DS. 2011. Gulf of Maine shells reveal changes in seawater temperature seasonality during the Medieval Climate Anomaly and the Little Ice Age. Palaeogeography, Palaeoclimatology, Palaeoecology, 302: 43–51.
Wanamaker Jr. AD, Kreutz KJ, Schöne BR, Pettigrew N, Borns HW, Introne DS, et al. 2008. Coupled North Atlantic slope water forcing on Gulf of Maine temperatures over the past millennium. Climate Dynamics, 31: 183–194.
Whitney NM, Johnson BJ, Dostie PT, Luzier K, and Wanamaker AD. 2019. Paired bulk organic and individual amino acid δ15N analyses of bivalve shell periostracum: A paleoceanographic proxy for water source variability and nitrogen cycling processes. Geochimica et Cosmochimica Acta, 254: 67–85,
Williams B. 2020. Proteinaceous corals as proxy archives of paleo-environmental change. Earth-Science Reviews, 209: 103326.
Wilson KL, Tittensor DP, Worm B, and Lotze HK. 2020. Incorporating climate change adaptation into marine protected area planning. Global Change Biology 26: 3251–3267.
Worm B, and Lotze HK. 2021. Marine biodiversity and climate change. In Climate and global change: observed impacts on Planet Earth, 3rd ed. Edited by T Letcher. Elsevier. pp. 445–464.
Yamamoto-Kawai M, Carmack E, and McLaughlin F. 2006. Nitrogen balance and Arctic throughflow. Nature 443: 43.
Supplementary material
Supplementary Material 1
- Download
- 17.85 KB
Supplementary Material 2
- Download
- 20.62 KB
Information & Authors
Information
Published In

FACETS
Volume 7 • Number 1 • January 2022
Pages: 1142 - 1184
Editor: Ryan Stanley
History
Received: 16 December 2021
Accepted: 10 June 2022
Version of record online: 18 August 2022
Notes
This paper is part of a collection titled “Climate change and the Canadian marine conservation framework”.
Copyright
© 2022 Lotze et al. This work is licensed under a Creative Commons Attribution 4.0 International License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Data Availability Statement
All relevant data are within the paper and in the Supplementary Material.
Key Words
Sections
Subjects
Plain Language Summary
Using the history of ocean and resource dynamics to foresee climate change
Authors
Author Contributions
HKL, SM, MB, KF, SDF, EG, BK, EO, NS, OS, ECS, and MK conceived and designed the study.
HKL, SM, JC, MB, MAD, LGS, LG, NG, SDK, MM, ITGP, KR, and ECS performed the experiments/collected the data.
HKL, JC, MB, MB, KF, EG, EO, WM, KR, OS, and MK analyzed and interpreted the data.
HKL, MB, KF, EO, OS, and MK contributed resources.
HKL, SM, JC, MB, KF, MAD, SDF, EG, LGS, LG, NG, BK, SDK, EO, MM, WM, ITGP, KR, NS, OS, ECS, and MK drafted or revised the manuscript.
Competing Interests
The authors have declared that no competing interests exist.
Metrics & Citations
Metrics
Other Metrics
Citations
Cite As
Heike K. Lotze, Stefanie Mellon, Jonathan Coyne, Matthew Betts, Meghan Burchell, Katja Fennel, Marisa A. Dusseault, Susanna D. Fuller, Eric Galbraith, Lina Garcia Suarez, Laura de Gelleke, Nina Golombek, Brianne Kelly, Sarah D. Kuehn, Eric Oliver, Megan MacKinnon, Wendy Muraoka, Ian T.G. Predham, Krysten Rutherford, Nancy Shackell, Owen Sherwood, Elizabeth C. Sibert, and Markus Kienast. 2022. Long-term ocean and resource dynamics in a hotspot of climate change. FACETS.
7: 1142-1184.
https://doi.org/10.1139/facets-2021-0197
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
Cited by
1. Public perceptions of aquaculture in Maine: The role of place-based values
2. Modeling embryo survival: Genetic and environmental influences on eye-up rate in a north American Atlantic salmon selective breeding program
3. Preliminary investigation of the local marine reservoir effect in Port Joli Harbour, Nova Scotia using archaeological M. arenaria shells
4. Mercury bioaccumulation in three colonial seabird species in the Gulf of Maine
5. Evaluating the influence of press and pulse disturbances on community dynamics of the Northeast US Large Marine Ecosystem
6. From land to deep sea: A continuum of cumulative human impacts on marine habitats in Atlantic Canada
7. Evidence of a Blue Crab (Callinectes sapidus) Successfully Overwintering in a Southern Maine Salt Marsh
8. Global research priorities for historical ecology to inform conservation
9. Using the Fossil Record to Understand Extinction Risk and Inform Marine Conservation in a Changing World
10. Habitat sensitivity in the West African coastal area: inferences and implications for regional adaptations to climate change and ocean acidification
11. Ocean Biomass and Climate Change
12. Uncertainty in the evolution of northwestern North Atlantic circulation leads to diverging biogeochemical projections
13. Impacts of Increasing Temperature on the Metabolism of Confined and Freely Moving American Lobsters (
Homarus americanus
)
14. Applying ensemble ecosystem model projections to future-proof marine conservation planning in the Northwest Atlantic Ocean
15. Chapter 21 : Northeast. Fifth National Climate Assessment