The effects of L-thyroxine and its association with physical exercise decrease the inflammation in an acute lung injury model
Abstract
Acute lung injury (ALI) is an inflammatory disease with a high incidence and prevalence that can lead to death. While L-thyroxine is widely used as a synthetic hormone in the treatment of hypothyroidism, its effects on inflammatory processes are not fully understood. Physical exercise, recommended for the general population, serves as an ally in treating various pathologies. Due to this and the lack of understanding of how L-thyroxine affects the immune response, coupled with the well-known benefits of exercise, this study aimed to evaluate L-thyroxine’s effects on lipopolysaccharide-induced inflammation and its combination with exercise in ALI. We assessed L-thyroxine’s effects on RAW 264.7 cells exposed to LPS and its association with exercise in a mouse ALI model. Pre-treatment with L-thyroxine demonstrated a decrease in the production of reactive oxygen species (P < 0.01) and the expression of pro-inflammatory genes and proteins (P < 0.05). In ALI, pre-treatment with L-thyroxine, either alone or combined with exercise, reduced the cell influx (P < 0.01) and the release of inflammatory cytokines (P < 0.05), in addition to mitigating lung damage caused by LPS (P < 0.01). However, combining L-thyroxine with exercise did not enhance its anti-inflammatory effect.
Introduction
Acute lung injury (ALI), a mild or moderate form of acute respiratory distress syndrome (ARDS), is characterized by intense inflammation and pulmonary edema. The clinical picture involves a reduction of lung compliance, dyspnea, and increased hypoxemia and ventilatory effort (Mowery et al. 2020; Saguil and Fargo 2020). In addition, the inflammatory cells that migrate into the lung generate changes in their functions and morphological features, complicating treatment (Grommes and Soehnlein 2011; Fei et al. 2019). For that reason, this disease is responsible for about 200 000 cases in the United States and has a high morbidity and mortality rate in critical patients. The cellular mechanism involved is related to the high production of inflammatory cytokines, e.g., tumor necrosis factor-alpha (TNF-α), interleukin-1beta (IL-1β), interleukin-6 (IL-6), interleukin-8, the activation and chemotaxis of macrophages and neutrophils, as well as the formation of free radicals, which assist in the persistence of the inflammatory picture (Robb et al. 2016; Mowery et al. 2020). It is considered an inflammation of difficult control and difficult diagnosis with little information about its development. However, ALI can progress during different phases, becoming a complex pulmonary syndrome with a broad spectrum (Lin et al. 2018).
Several predictive models have been used to assess the severity and outcomes of ALI, as well as its complications. Among them, the Acute Physiology and Chronic Health Evaluation score considers acute and chronic physiological variables to predict mortality, and the Sequential Organ Failure Assessment score evaluates organ dysfunction and is a reliable predictor of mortality (Knaus et al. 1985; Vincent et al. 1996). Additionally, the Lung Injury Prediction Score is used to predict the risk of developing ARDS in hospitalized patients, while the Berlin definition categorizes ARDS severity based on clinical and gasometric criteria (Ranieri et al. 2012). Common complications associated with ALI include respiratory failure, multiple organ dysfunction, and secondary infections, which are efficiently predicted by these models (Gajic et al. 2011; Ranieri et al. 2012). Furthermore, individuals recovering from ALI may experience long-term physical and mental health impairments that affect their quality of life (Fan et al. 2014; Huang et al. 2016). Readmissions may be necessary, and hospital costs per person are estimated to range from $8476 to $547 974 (Boucher et al. 2022). Therefore, its prevention and the search for new therapeutical approaches become relevant strategies.
Thyroid hormones (TH) are crucial for individuals’ development and physiological metabolism, also causing severe pathologies when thyroid dysfunction occurs (Mullur et al. 2014). L-thyroxine is considered a gold standard in treating hypothyroidism. In addition to affecting life quality, the hypothyroidism can also cause an inflammatory clinical picture that seems to be positively modulated by this hormone (Tellechea 2021). In an experimental model, a treatment performed with L-thyroxine in rats with hypothyroidism showed a reduction in inflammatory mediators such as C-reactive protein, IL-6, and TNF-α (Abbas and Sakr 2016). Other evidence with L-thyroxine demonstrated modulation in the proliferation of macrophages, migration, senescence, and secretion of inflammatory factors, in addition to improvements in memory deficits and oxidative stress (Ning et al. 2018; Bavarsad et al. 2020).
There are inflammatory aspects common to both hypothyroidism and ALI. Hypothyroidism is characterized by elevated levels of inflammatory cytokines, factor also present in ALI. L-thyroxine may potentially exert a beneficial anti-inflammatory effect in ALI, by normalizing the elevated cytokine levels, as it does in hypothyroidism. Besides, the symptoms and complications of hypothyroidism, such as fatigue, cardiovascular diseases, and musculoskeletal issues (Magri et al. 2010; Chaker et al. 2017), are conditions that patients with ALI, especially those in intensive care units for extended periods, may also experience. Therefore, we decided to investigate whether pre-treatment with L-thyroxine, a widely used medication, could reduce inflammatory levels in ALI.
Additionally, the physical exercise (Ex) is a viable and widely recommended option to assist drug treatment. This natural resource has shown benefits in the prevention and adjuvant treatment for individuals with various diseases and age ranges (Booth et al. 2012; Sáez de Asteasu et al. 2019; Martha et al. 2021). Chronic aerobic exercise has been extensively studied and proven to be effective in improving overall health and enhancing the immune system. It is known to induce beneficial adaptations, including enhancing the body’s antioxidant capacity, reducing the production of pro-inflammatory cytokines, and increasing the production of anti-inflammatory cytokines (Bruunsgaard 2005; Gleeson et al. 2011). These changes are crucial for modulating inflammation and preparing the body to combat severe inflammatory conditions, such as ALI. Also, Ex has been increasingly studied, especially in experimental models for combating ALI (Gholamnezhad et al. 2022) and sepsis (Wang et al. 2021).
From this perspective, this study aimed to evaluate the effects of L-thyroxine in Raw 264.7 macrophages stimulated by lipopolysaccharide (LPS) and their association with Ex in a mouse model of ALI. Considering that L-thyroxine is an essential hormone to maintain body homeostasis and that Ex acts systemically in the organism, we hypothesized that pre-treatment with L-thyroxine and its association with Ex could mitigate the inflammation caused by LPS, modulating the inflammatory response.
Materials and methods
In vitro cell culture
The RAW 264.7 (CLS Cat# 400319/p462, RRID:CVCL_0493) cell line from the ATCC bank (Virginia, USA) was cultivated in Eagle medium modified by Dulbecco (DMEM; GibcoTM, Life-technologies, USA) medium and supplemented with 10% fetal bovine serum (FBS; GibcoTM, Life-technologies, USA) and the antibiotics penicillin and streptomycin (10 000 units/mL, GibcoTM, Life-technologies, USA) at 50 units/mL at 37 °C in an incubator moistened with 5% CO2. All experiments were performed in triplicate and only cells that did not exceed 12 passages were used.
Preparation and pre-treatment with L-thyroxine
The stock solution of L-thyroxine (Sigma–Aldrich, USA) was dissolved in dimethyl sulfoxide (DMSO; Merck, USA) and then diluted in DMEM at different concentrations (50 nmol/L- 150 nmol/L). These concentrations were used according to Ning et al. (2018). The RAW 264.7 cells were distributed into 6–24-well plates according to each experiment. After 24 h, the cells were pre-treated with these different concentrations of L-thyroxine for 1 h. Subsequently, the cells were activated with 1 µg/mL of LPS (Escherichia coli 026: B6, Sigma–Aldrich, USA) for 12 h. Next, the cells were collected for the analyses described below, which assisted in the choice of the therapeutical and study concentration of L-thyroxine. Dexamethasone (Dex; Sigma–Aldrich, USA) was used as the positive control in the tests (10 µmol/L).
Cell viability assay
The effect of L-thyroxine on cell viability was evaluated through the bromide of 3-[4,5- dimethyl thiazole -2-il]-2,5 diphenyl tetrazolium (MTT; Sigma–Aldrich, USA) assay. The cells were distributed into 24-well plates containing 0.6 mL of DMEM. MTT was added into all plate wells and the plates were incubated for 3 h. Next, the supernatant was discarded and DMSO was added to dissolve the formazan crystals produced by MTT. The optical density was read in an ELISA microplate reader (EZ Read 400; Biochrom, USA) at the wavelengths of 570/620 nm.
Antioxidant activity
The 2,2-diphenyl-1-picrylhydrazyl (DPPH; Sigma–Aldrich, USA) method was used to determine the antioxidant activity percentage of L-thyroxine. DPPH is a purple, stable free radical that can be reduced in the presence of an antioxidant molecule, giving origin to a colorless solution. Ascorbic acid (AsA; 550 µg/mL; Sigma–Aldrich, USA) was used as a positive control. The optical density was measured at 515 nm using an ELISA microplate reader (Kedare and Singh 2011).
Evaluation of the reactive oxygen species
The diacetate of 2′,7′- dichlorofluorescein (DCFH-DA; Sigma–Aldrich, USA) assay was performed to evaluate the generation of intracellular reactive oxygen species (ROS). The DCFH-DA is a non-fluorescent compound that, when oxidated by ROS, becomes fluorescent and gives origin to 2′,7′- dichlorofluorescein (DCF; Sigma–Aldrich, USA) (Kalyanaraman et al. 2012). Next, 1 × 105 cells were cultivated and treated with L-thyroxine (150 nmol/L), hydrogen peroxide (100 umol/L) or LPS (1 ug/mL). Ascorbic acid (550 µg/mL) was used as an antioxidant control. Later, cells were washed twice with phosphate buffer saline (PBS) and incubated with 10 µmol/L of DCFH-DA at 37 °C for 30 min. The fluorescence intensity was measured by VICTOR® microplate reader (PerkinElmer, USA) at an excitation wavelength of 485 nm and at an emission wavelength of 520 nm.
Western blot
The cells were seeded in 6-well microplates and then lysed with a specific buffer containing protease inhibitors. The total protein concentration in the samples was measured with a NanoDrop Lite spectrophotometer (Thermo Scientific, USA). Aliquots from each sample contained equal amounts of protein (50 µg), which ran in 10% polyacrylamide gel (SDS-page) and were transferred to a nitrocellulose membrane (California, USA). Soon after, the membranes were blocked with a buffered saline solution and skim powdered milk for 1 h. Next, the material was incubated overnight at 4 °C with the primary antibodies anti-NFκB p-p65 (1:250, Cell Signaling Technology Cat# 3033, RRID:AB_331284) anti-COX-2 (1:500, Cell Signaling Technology Cat# 12282, RRID:AB_2571729) and anti-GAPDH (1:1000, Thermo Fisher Scientific Cat# 39-8600, RRID:AB_2533438). Subsequently, the blots were incubated with the secondary antibody anti-IgG (1:2000, Thermo Fisher Scientific, Waltham, USA) for 2 h. The protein bands were visualized by enhanced chemiluminescence (GE Lifescience). Band quantification was performed using the software ImageJ.
Gene expression
The cells were distributed into 6-well microplates containing 2 mL of DMEM, and the total RNA of the cell culture was obtained using Trizol (Invitrogen- Massachusetts, USA) according to the manufacturer’s recommendations. Next, cDNA was synthesized through reverse transcription using a commercial kit GoscriptTM Reverse Transcriptase (Promega, USA) with 5 µg of total RNA for each sample. The real-time (RT-qPCR) reactions were performed using an SYBR Green PCR Master Mix (Applied Biosystems; Thermo-Fisher Scientific, USA) to identify the synthesis of double strands in the reaction. Beta-2-microglobulin (B2M) was used as an endogenous control. The relative expression of mRNA was calculated using the ΔΔCq method (Livak and Schmittgen 2001). The sequence of primers used was: B2M, forward, 5′-CCCCAGTGAGACTGATACATACG-3′ and reverse, 5′-CGATCCCAGTAGACGGTCTTG-3′; TNF-α, forward, 5′-ATAGCTCCCAGAAAAGCAAGC-3′ and reverse, 5′-CACCCCGAAGTTCAGTAGACA-3′; IL-10, forward, 5′-GCCAAGCCTTATCGGAAATG-3′ and reverse, 5′-AAATCACTCTTCACCTGCTCC-3′; IL-6, forward, 5′-TGGAGTCACAGAAGGAGTGGCTAAG-3′ and reverse, 5′-CTGACCACAGTGAGGAATGTCCAC-3′. RT-qPCR was performed with an initial step of 20 s at 95 °C, followed by 40 cycles with 10 s at 95 °C and 30 s at 60 °C. The melting curve profile of the PCR products was obtained after an initial step of 15 s at 95 °C, followed by an incubation at 60 °C for 1 min (for reannealing) and heating of 0.3 °C/s to 95 °C.
In vivo experiments in animals
The study was carried out with male C57BL/6 mice (RRID: MGI:2159769). All studied animals were between 8 and 12 weeks old, weighed between 23 and 25 g and were provided by the Center for Experimental Biological Models of the Pontifical Catholic University of Rio Grande do Sul (PUCRS). To achieve a more precise control of variables, it was decided to use male mice. Hormonal levels can influence the inflammatory response, and male mice tend to have more stable hormonal levels compared to females, which undergo reproductive cycles. The animals were split into groups of five per cage (22 cm × 16 cm × 14 cm), over shelves at a controlled temperature (24 ± 2 °C), a light/dark cycle of 12 h, with free access to food and water, and were acclimated for a week before the study. The experimental protocol was approved by the Ethics Committee on Animal Experimentation of PUCRS (number 9611). All experiments were conducted following the National Institutes of Health Guide for the Care and Use of Laboratory Animals.
Methodological approach and ALI induction
The animals were split and handled according to their experimental groups as described below:
1.
Control: The mice were anesthetized by isoflurane inhalation and intranasal (IN) PBS was administered. Next, DMSO was administered via the intraperitoneal (IP) route.
2.
ALI: The mice were anesthetized by isoflurane inhalation and received LPS via the IN route (2 mg/kg) to induce ALI (Haute et al. 2023).
3.
ALI + Dex: The mice were anesthetized by isoflurane inhalation and received LPS via the IN route. Next, Dex (1 mg/kg) was administered via the IP route (Tu et al. 2020).
4.
ALI + L-thyroxine: The mice were anesthetized by isoflurane inhalation and received LPS via the IN route. Next, L-thyroxine (30 µg/kg) was administered via the IP route. The concentration chosen was based on the proportion of the animal’s weight, according to previous studies with L-thyroxine and the treatment time adapted to the ALI induction protocol (Seyedhosseini Tamijani et al. 2019; Bavarsad et al. 2020).
5.
Ex + ALI: The mice performed Ex for five weeks and were then anesthetized by isoflurane inhalation and administration of LPS via the IN route.
6.
Ex + ALI + Dex: The mice performed Ex for 5 weeks and were then anesthetized by isoflurane inhalation and IN administration of LPS. Next, dexamethasone was administered via the IP route.
7.
Ex + ALI + L-thyroxine: The mice performed Ex for five weeks and were then anesthetized by isoflurane inhalation and IN administration of LPS. Next, L-thyroxine was administered via the IP route.
The mice were anesthetized with ketamine and xylazine (0.4 and 0.2 mg/g, respectively) 12 h after ALI induction and euthanized by exsanguination through cardiac puncture. Next, the bronchoalveolar lavage (BAL) and lung tissue samples were collected.
Exercise
Animals from groups Ex + ALI; Ex + ALI + Dex; and Ex + ALI + L-thyroxine were submitted daily to an exercise session on a motorized treadmill, during the 5 weeks that preceded the day of ALI induction. The animals were familiarized with the treadmill in 10 min sessions at a speed of 5 m/min for one week to reduce stimuli related to handling and the environment. Initially, the brisk walk speed and time were 10 m/min, being 20 min/day on the first day with an increment of 10 min/day until reaching 60 min/day, to reach an exercise intensity of approximately 70% of maximum oxygen consumption (VO2) of the animals. After that, exercise sessions at a speed of 10 m/min for 60 min, 5 days a week, were used (Wu et al. 2007). Before starting the protocol, the animals were habituated to the room for 30 min. No stimulus, such as an electric shock, was applied to motivate the animals to brisk walk. Animals that refused to brisk walk, presented any visual sign of distress, or had any difficulty to follow the protocol were exclude from the study.
Bronchoalveolar lavage sampling
The animals were previously anesthetized (0.4 mg/g ketamine and 0.2 mg/g xylazine), and BAL sampling was performed by applying an injection and aspiration of 1 mL of PBS containing 2% SFB (process performed twice) using a tracheal cannula initially placed into the animal.
Total and differential cell count
The BAL material collected was centrifuged (1000 × g for 10 min) and the supernatant was reserved. The cell pellet was diluted with 350 µL of PBS to perform the counts. The exclusion method with Trypan blue in a Neubauer chamber (Boeco, Germany) allowed performing the total cell count and calculating cell viability. Slides were prepared to evaluate the differential cytology of inflammatory cells using 80 µL of cell material previously diluted with PBS in a centrifuge (FANEM, São Paulo, Mod. 218) at 500 rpm for 5 min. Next, the slides were fixed in ambient air and stained with hematoxylin and eosin (H&E) (Panótico Rápido; Laborclin, Brazil). The different cell types were analyzed by optical microscope and, after counting 400 cells, the relative and/or absolute numbers were expressed.
Cytokine quantification
The lung tissue samples were collected and macerated with a PBS solution. The simultaneous quantification of cytokines IL-1β, TNF-α, IL-6, and IL-10 was performed using ProcartaPlex (Thermo Fisher Scientific, USA) in the MagPix device (MILLIPLEX®), and the software xPONENT® 4.2 (MILLIPLEX®) was used to analyze the data generated.
RNA and RT-qPCR extraction from lung tissue
After euthanasia, the lung tissue was separated and stored in an ultra-freezer (−80 °C) for RNA extraction. The total RNA was extracted from all samples using Trizol, following the manufacturer’s recommendations. cDNA synthesis and RT-qPCR were performed as mentioned in the “Gene expression section” section, using the same primers and methods.
Histopathological analysis of lung tissue
For this procedure, 10% formaldehyde was used in the lung tissues, after which they were stored for 24 h in formalin. Next, the tissues were embedded with liquid paraffin, and, after that, the slides were produced with 4 µm thick histological sections. Later, H&E staining was performed to identify the inflammatory infiltrate. The changes in lung tissues were assessed through a semiquantitative evaluation approach (Zhang et al. 2017). In summary, histological criteria were blindly assigned to measure the severity of pulmonary inflammation, including interstitial inflammation, infiltration of inflammatory cells, congestion, and edema. The histopathological assessments ranged from 0 (normal state) to 4 (severe grade) and were determined by averaging the assessments assigned to each individual mouse in their respective groups. The images were captured with a DP73 camera coupled to a light microscope BX43 (Olympus, Japan).
Statistical analysis
At first, we conducted in vitro experiments with RAW 264.7 cells, performing three independent experiments in triplicates to evaluate whether our hypothesis about the ability of L-thyroxine to reduce inflammation was warranted or not. Then, we presented the results obtained with the animals (in vivo) as mean ± standard deviation (SD). We used five animals per group with the aim of minimizing the need for animals in research, in strict adherence to ethical considerations that promote the right to life and animal welfare, while ensuring the validity of the results. Studies with similar approaches have also adopted a comparable sample size (Scheffer et al. 2019; Jin et al. 2020). Data were expressed as mean ± SD and were analyzed by one-way Analysis of variance (ANOVA). P < 0.05 was defined and considered statistically significant. Subsequently, Tukey’s post-test for multiple comparisons was performed. All statistical analyzes were performed using GraphPad Prism 8 software (GraphPad Software, CA).
Results
In vitro
L-thyroxine reduces the proliferation of RAW 264.7 cells activated by LPS
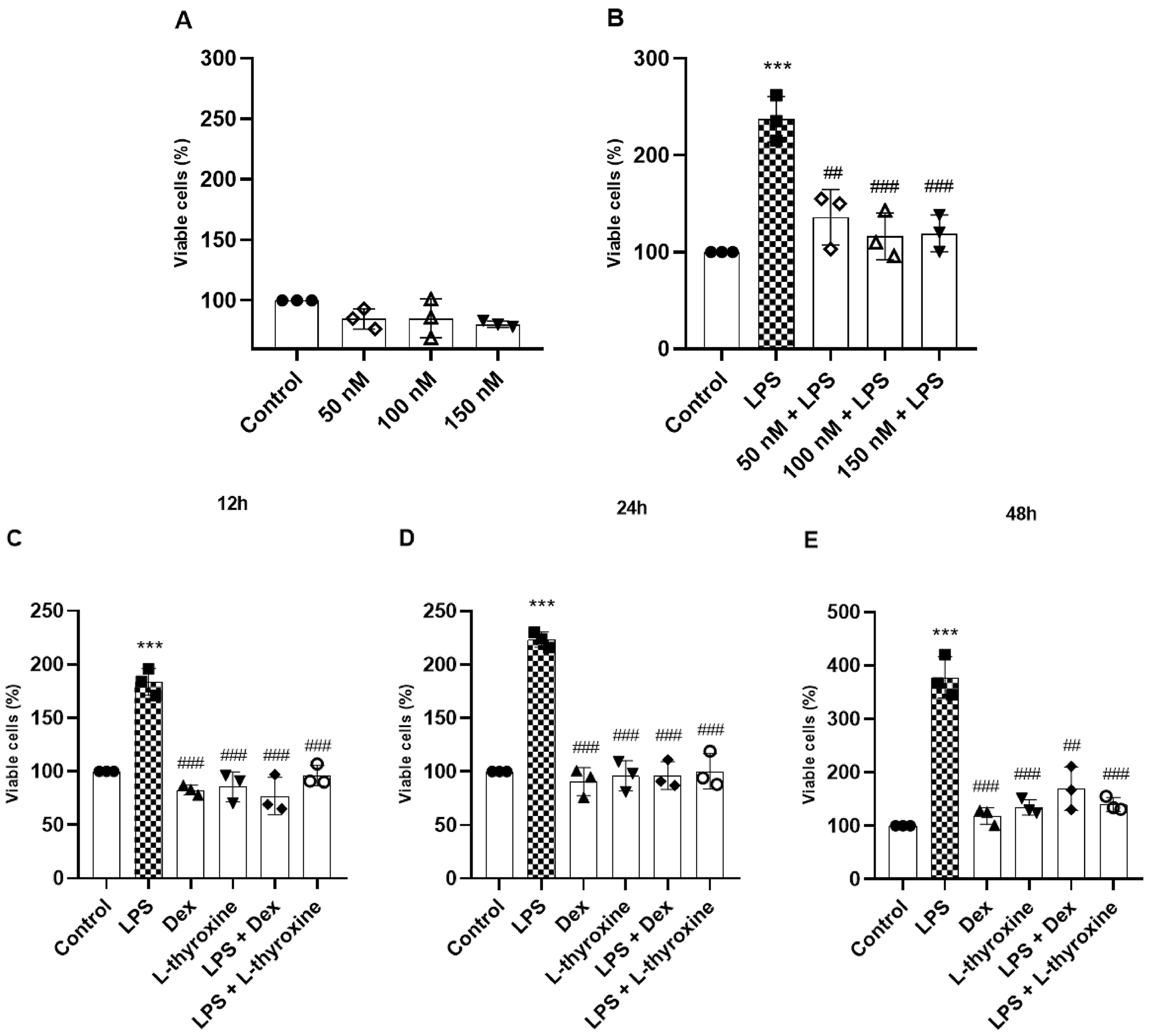
Our investigation started by evaluating the cell viability of RAW 264.7 cells after 24 h of pre-treatment with L-thyroxine (Fig. 1a). There was no significant reduction in viable cells at any of the concentrations tested, indicating low cytotoxicity. Next, we evaluated the effects of L-thyroxine in the proliferation of cells stimulated with LPS. The RAW 264.7 cells were pre-treated for 1 h with L-thyroxine at the same concentrations tested, followed by LPS stimulation for 24 h. LPS significantly increased cell proliferation, and all L-thyroxine concentrations reduced this response (Fig. 1b). With that, we chose the L-thyroxine concentration of 150 nmol/L for the rest of the study, observing its effects at different times (Figs. 1c–1e) and including a positive control with an established anti-inflammatory effect, dexamethasone. L-thyroxine and dexamethasone reduced, at all times analyzed, the activation of cell proliferation induced by LPS.
Fig. 1.
L-thyroxine decreases the production of ROS in RAW 264.7 cells
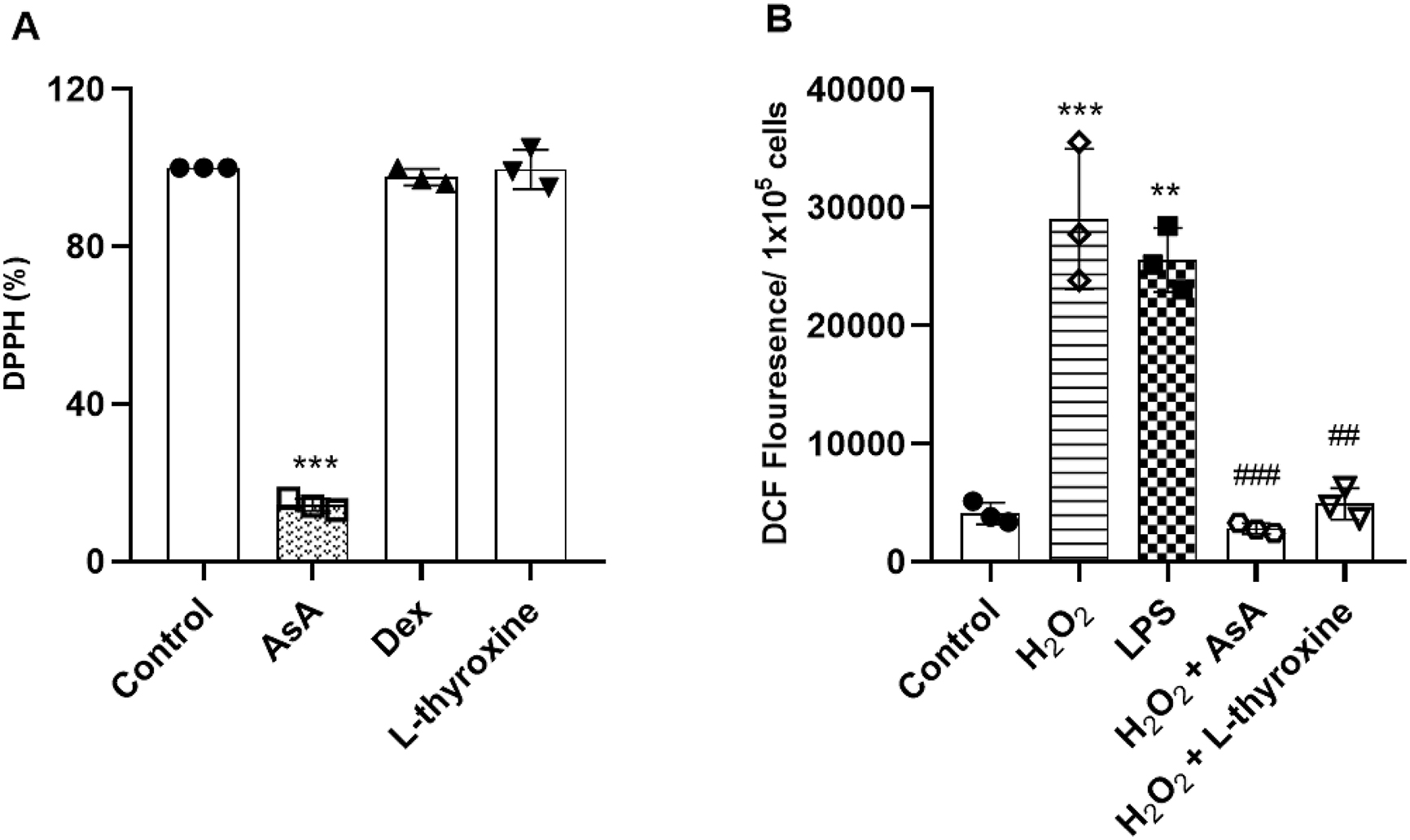
To evaluate the antioxidant potential of L-thyroxine, we used the DPPH assay. Our results did not show the antioxidant effects of L-thyroxine, and the same occurred for dexamethasone (Fig. 2a).ROS are an important factor for oxidative stress, very present in ALI. Therefore, we evaluated the generation of ROS in RAW 264.7 cells through the DCFH-DA method. Cells stimulated with hydrogen peroxide (100 µmol/L) and LPS (1 µg/mL) increased the ROS, and cells pre-treated with L-thyroxine (150 nmol/L) and dexamethasone (10 µmol/L) reduced this effect (Fig. 2b).
Fig. 2.
L-thyroxine regulates the protein expression of Cox-2 and p-p65
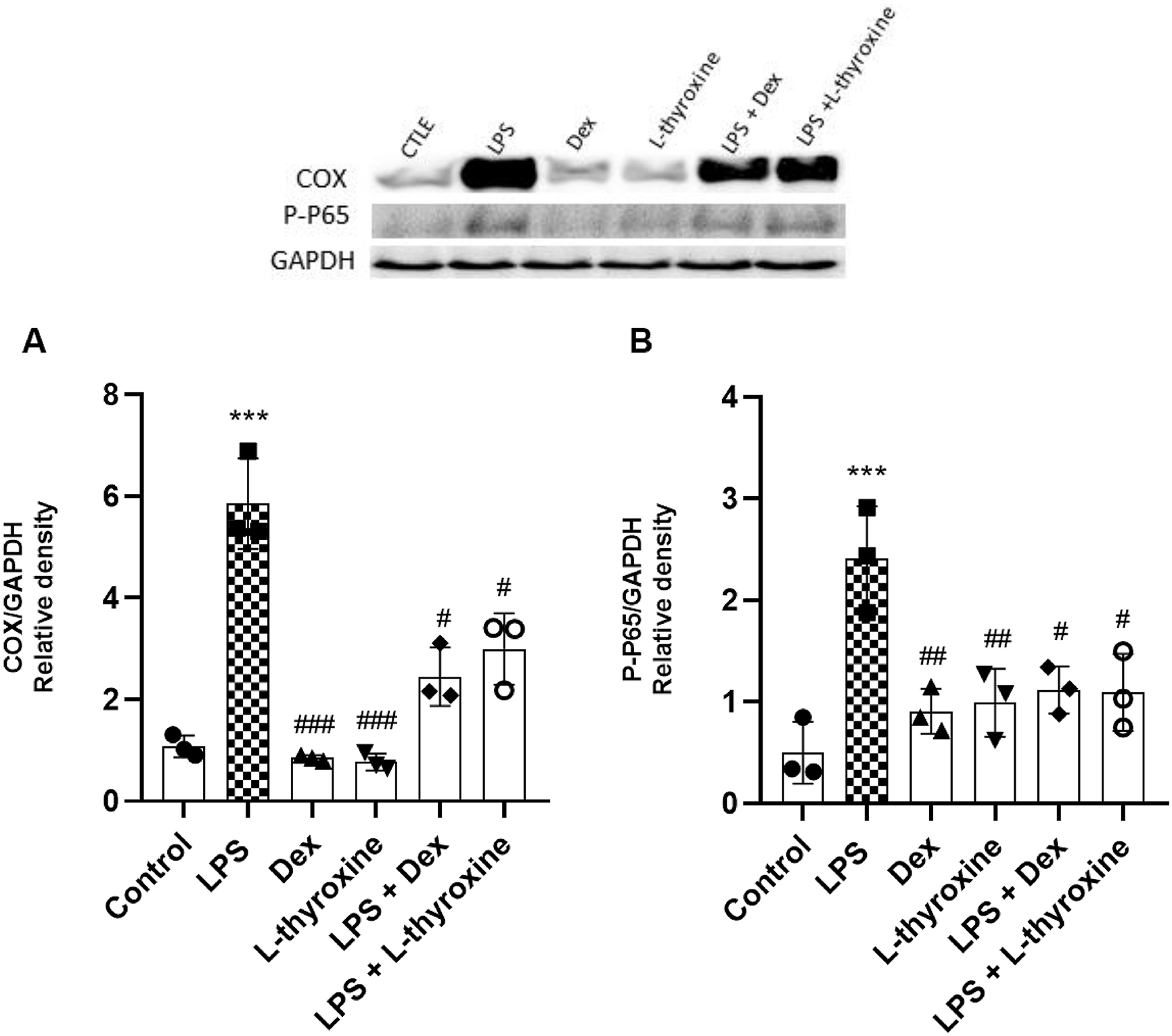
Through Western Blot, we evaluated the expressions of important proteins in the inflammatory process: Cox-2 and phosphorylated NF-kB (p-p65). The cells were pre-treated with L-thyroxine (150 nmol/L) and Dex (10 µmol/L) for 1 h and then exposed to LPS (1 µg/mL). Our data have shown that LPS induced an increase in the expression of Cox-2 and p-p65 compared to the control group, whereas pre-treatment with L-thyroxine and dexamethasone reduced these expression levels compared to the LPS group (Figs. 3a and 3b).
Fig. 3.
L-thyroxine modulates the gene expression of IL-6, TNF-α, and IL-10 in RAW 264.7 cells stimulated by LPS
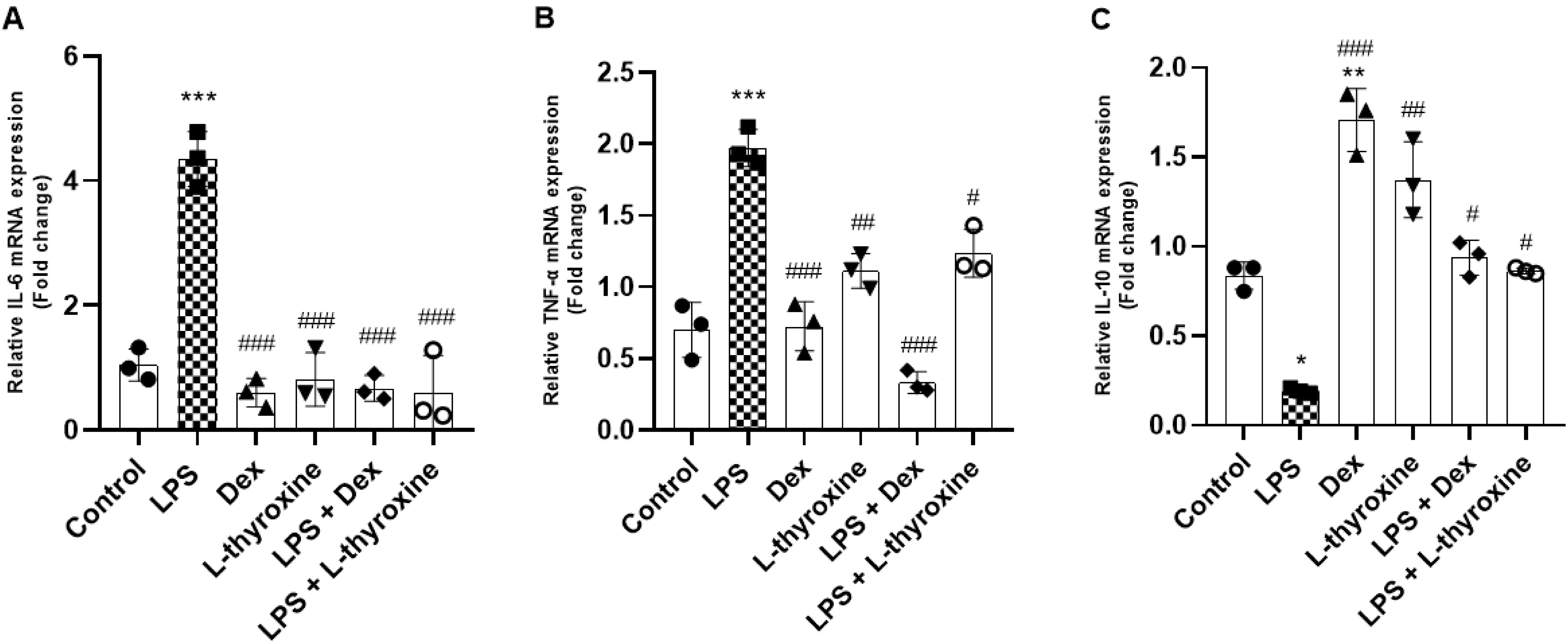
To evaluate the effects of L-thyroxine on the expression of pro- and anti-inflammatory genes, the mRNA levels of IL-6, TNF-α, and IL-10 were evaluated by real-time PCR. The cells were pre-treated with L-thyroxine (150 nmol/L) and Dex (10 µmol/L) for 1 h and then exposed to LPS (1 µg/mL). The LPS group significantly increased the mRNA levels of IL-6 and TNF-α, and pre-treatment with L-thyroxine, as well as treatment with dexamethasone, mitigated the gene expression of these genes caused by LPS (Figs. 4a and 4b). Furthermore, although pre-treatment with L-thyroxine did not increase the expression of mRNA of IL-10 compared to the control group, as seen in the group pre-treated with dexamethasone only, he it was capable of preventing the drop in the mRNA levels of IL-10. The same was observed in the groups pre-treated with dexamethasone and in the isolated L-thyroxine group (Fig. 4c).
Fig. 4.
In vivo
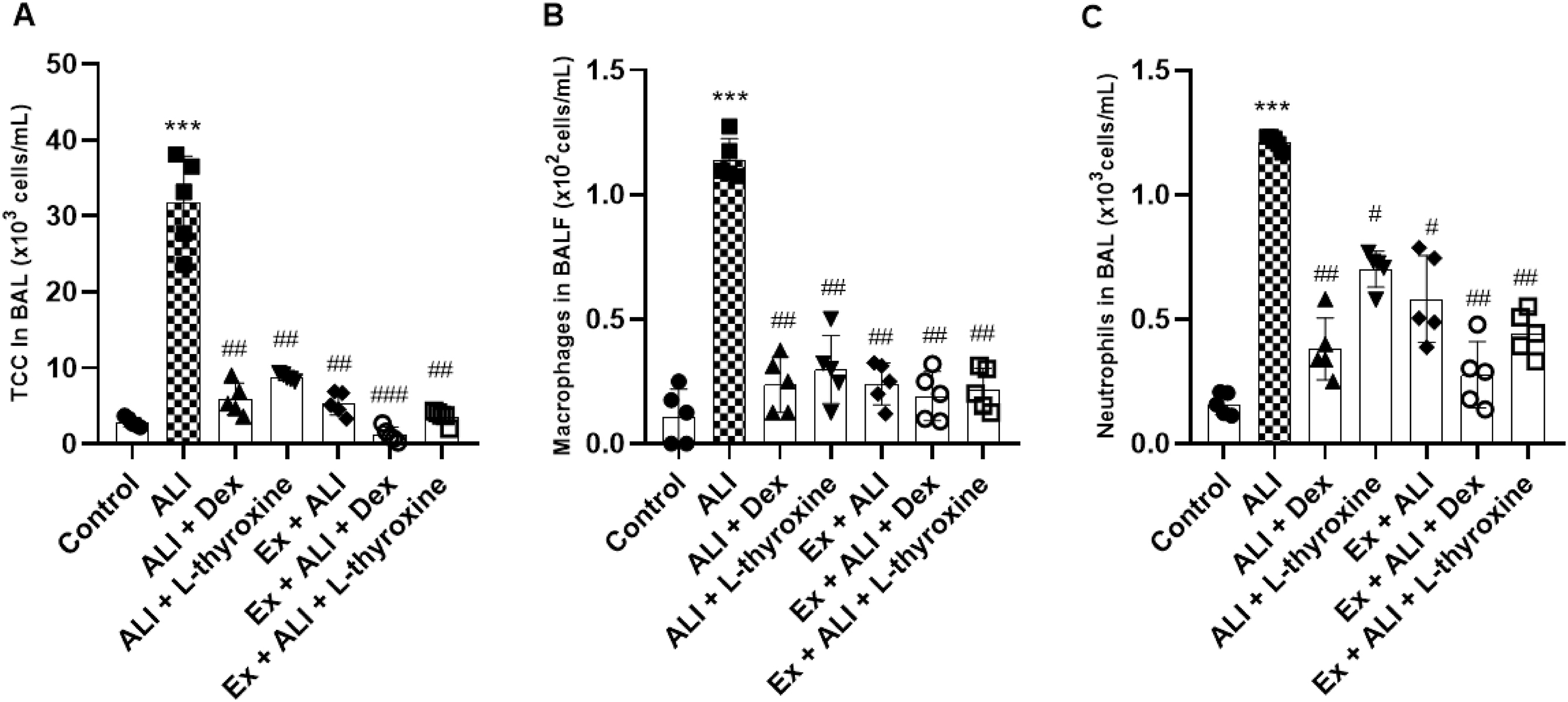
L-thyroxine and physical exercise decrease the total and differential cell count in the BAL of mice with ALI induced by LPS
To determine whether L-thyroxine and its association with Ex could regulate the influx in inflammatory cells, we performed the total and differential cell count from the BAL of mice. ALI induced by LPS significantly increased the migration of macrophages and neutrophils in the BAL of the animals in the total and differential counts. However, pre-treatments with L-thyroxine, dexamethasone, and the association with Ex decreased the displacement of these inflammatory cells into the lungs of the mice (Figs. 5a–5c).
Fig. 5.
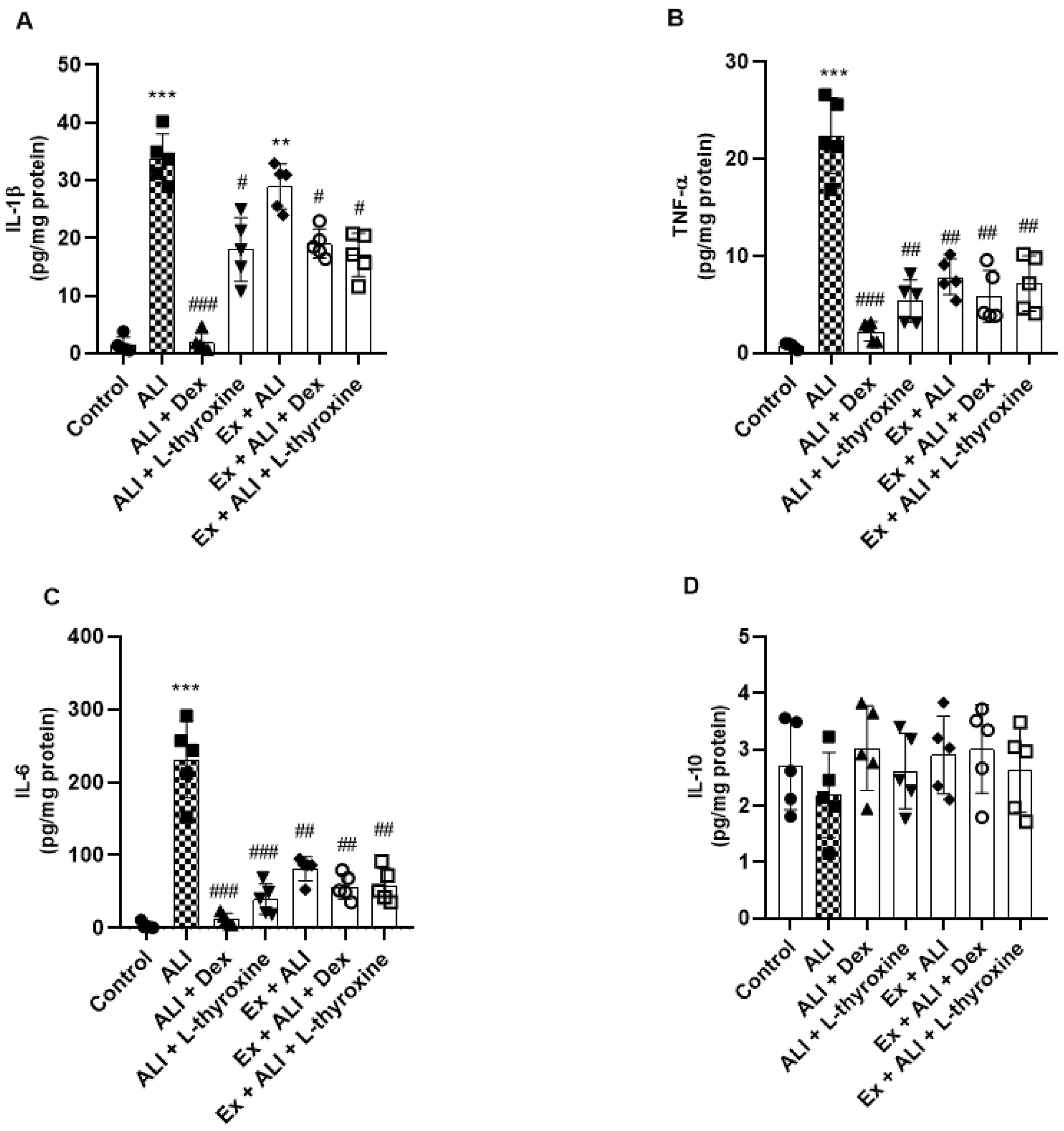
L-thyroxine and physical exercise reduce the release of inflammatory cytokines
We also investigated the levels of IL-1β, TNF-α, IL-6, and IL-10 in the lung tissue of mice using a ProcartaPlex assay kit. The animals with ALI showed an increase in the IL-1β compared to the control group, whereas pre-treatments with L-thyroxine, dexamethasone, and its association with Ex reduced this increase. Furthermore, it was observed that observed that physical exercise did not reduce the levels of IL-1β (Fig. 6a). With regard to cytokines IL-6 and TNF-α, our findings showed a significant increase in the ALI group and a reduction in the groups with L-thyroxine, dexamethasone, and the associations with Ex (Figs. 6b and 6c). No significant differences were found in the production of IL-10 (Fig. 6d).
Fig. 6.
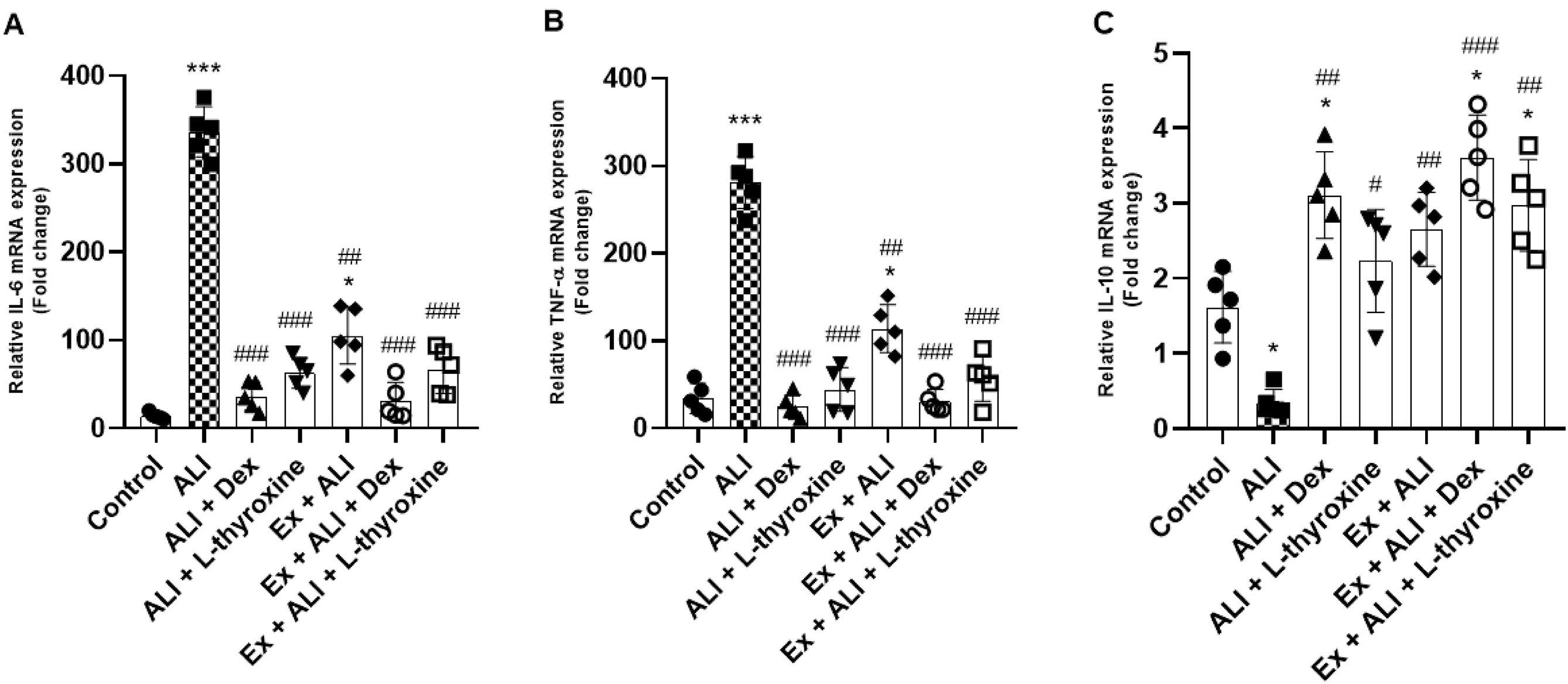
L-thyroxine and physical exercise modulate the gene expression of cytokines in mice with ALI
We verify below the mRNA of genes IL-6, TNF-α, and IL-10 in the lungs of the studied animals by real-time PCR. The results showed a significant increase in the gene expression of IL-6 and TNF-α in animals with ALI induced by LPS, whereas pre-treatment with L-thyroxine, dexamethasone, and the other groups with Ex reduced this expression. Nevertheless, we observed that Ex did not prevent the increase of these cytokines compared to the control group. However, the increase of these pro-inflammatory genes showed a significant decrease compared to mice with ALI (Figs. 7a and 7b), indicating a protective effect, nonetheless. The mRNA levels of IL-10 were reduced in the ALI group, and pre-treatment with dexamethasone increased them. A similar effect was demonstrated in the pre-treatments with L-thyroxine and dexamethasone associated with Ex. Although pre-treatment with L-thyroxine did not show an increase in IL-10 mRNA expression, it was able to inhibit its decrease in animals with ALI, as well as Ex (Fig. 7c).
Fig. 7.
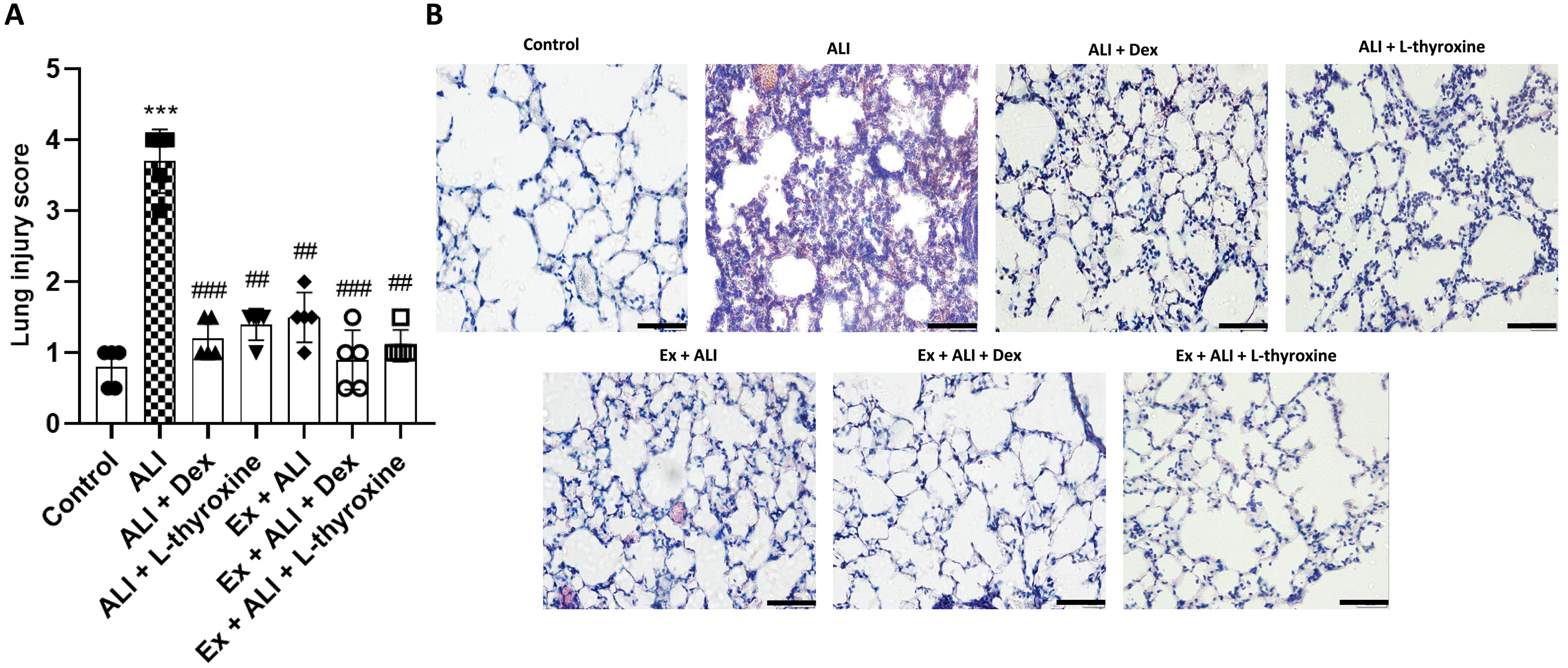
L-thyroxine and physical exercise reduce histopathological changes in the lung of mice with ALI induced by LPS
Finally, we analyzed whether pre-treatment with L-thyroxine and its association with Ex would be able to reduce the migration of inflammatory cells into the lungs of mice with ALI. As observed in Fig. 8a, mice with ALI presented a increase in the lung injury score, indicating the influx of macrophages and neutrophils, and consequent changes in lung tissue structures. On the other hand, pre-treatments with L-thyroxine, dexamethasone, and associations with Ex mitigated this effect caused by LPS. To observe these changes, we stained lung tissue sections with H&E (Fig. 8b).
Fig. 8.
Discussion
Our results demonstrated that pre-treatment with L-thyroxine positively regulated the inflammatory response in macrophages stimulated by LPS. Since the lung is one of the most affected organs by the release of pro-inflammatory mediators during sepsis (Winters et al. 2010) and trauma (Pierce and Pittet 2014), we decided to investigate whether L-thyroxine could cause a protective effect in an ALI model induced by LPS. Furthermore, we included Ex as it is recommended to assist in treating hypothyroidism symptoms (Werneck et al. 2018) and there is evidence of its beneficial effects in ALI (Gholamnezhad et al. 2022). Thus, we evaluated the effects of L-thyroxine in association with Ex on the inflammatory levels of mice with ALI. Although the anti-inflammatory effects shown by L-thyroxine were not summed to those of Ex, our results indicate that combining L-thyroxine with Ex was beneficial against the lung inflammation caused by LPS. Therefore, we demonstrated, for the first time, the capacity of L-thyroxine in modulating mechanisms related to ALI and its effects in association with Ex in ALI induced by LPS.
The RAW 264.7 cell line has been widely used to investigate in vitro inflammatory function, particularly due to its response to LPS, which promotes the release of inflammatory cytokines such as IL-6 and TNF-α (Tang et al. 2019; Yi et al. 2019). Our results corroborate these findings, demonstrating that exposure to LPS significantly increased the levels of these cytokines. Additionally, we observed that LPS stimulates the generation of ROS, consistent with literature evidence (Baek et al. 2020). We found that L-thyroxine, although not demonstrating antioxidant activity via the DPPH method, reduced ROS formation in RAW 264.7 cells exposed to H2O2. These findings are supported by studies indicating that hypothyroidism increases ROS production (Santi et al. 2010; Cheserek et al. 2015) and L-thyroxine treatment normalizes this production (Chakrabarti et al. 2016).
Furthermore, L-thyroxine modulated the inflammatory response by reducing the expression of Cox-2 and the phosphorylated subunit of NF-kB, p-p65, which are crucial regulators in the inflammatory process. Selective Cox-2 inhibition is a well-established mechanism of action for non-steroidal anti-inflammatory drugs (Zarghi and Arfaei 2011) and NF-kB activation is a fundamental pathway in inflammatory response (Liu et al. 2017). Our results indicate that L-thyroxine may exert a significant anti-inflammatory effect by interfering with these mechanisms.
As described by Qin et al. (2016), LPS exposure elevates mRNA expression of IL-6 and TNF-α in RAW 264.7 cells. In our study, L-thyroxine demonstrated the ability to reduce these increases. Interestingly, L-thyroxine did not increase IL-10 mRNA expression but prevented its decline induced by LPS. This may be attributed to the action of TH, including L-thyroxine, in maintaining organismal homeostasis (McAninch and Bianco 2014) and modulating immune responses (De Vito et al. 2012).
In the literature, it is documented that inflammatory cytokines and C-reactive protein are elevated in hypothyroidism (Tayde et al. 2017; Tellechea 2021) and L-thyroxine treatment normalizes these levels (Marfella et al. 2011), indicating a positive role in stabilizing the immune response. Experimental models show anti-inflammatory actions of TH and their receptors (Chen et al. 2012; Furuya et al. 2017). In critically ill patients with sepsis, low levels of circulating L-thyroxine are found in plasma (Al-Abed et al. 2011), yet the role of TH in innate immune response remains poorly understood, with conflicting results (Wenzek et al. 2022). In our ALI model, L-thyroxine, both alone and in combination with Ex, modulated the inflammatory response, decreased inflammatory mediators, and improved tissue function, which are significantly altered in ALI.
Previous studies indicate that exercise can alter the profile of pro and anti-inflammatory cytokines depending on its execution protocol (Improta-Caria et al. 2021). In intensive care units, aerobic exercise on a cycle ergometer as early mobilization has been used to prevent delirium, reduce muscle loss, and improve lung function in critically ill patients, including those with ALI (Aquim et al. 2019). These interventions reinforce the idea that physical exercise is not only safe but also beneficial in critical health contexts. Furthermore, the American Heart Association has published guidelines for primary prevention of cardiovascular diseases, recommending at least 150 min per week of moderate-intensity physical activity or 75 min per week of vigorous-intensity exercise (Arnett et al. 2019), further encouraging regular exercise practice.
Moderate-intensity aerobic exercise on a treadmill has been shown to decrease IL-6 and TNF-α levels in both humans (Abd El-Kader et al. 2013) and mice (Lang et al. 2020) and our results confirm that moderate exercise can reduce IL-6 and TNF-α levels in mice. Additionally, we observed an increase in IL-1β, IL-6, and TNF-α levels in animals with ALI, which was significantly reduced with L-thyroxine pretreatment and in combination with exercise. Previous studies have identified elevated levels of IL-6, TNF-α, and IL-1β associated with a worse prognosis in ALI (Meduri et al. 1995; Voiriot et al. 2017) and our findings corroborate the increase in these inflammatory agents in ALI. Gene expression also reflected these changes, with a reduction in IL-6 and TNF-α mRNA levels following pretreatments.
While we did not observe a significant difference in IL-10 through the ProcartaPlex assay, the combination of L-thyroxine with exercise increased IL-10 mRNA expression. This synergy is crucial, as both L-thyroxine and exercise alone were able to prevent IL-10 levels from declining but not to increase them. The ability of exercise to increase IL-10 levels (Abd El-Kader and Al-Shreef 2018), combined with the effects of L-thyroxine, contributes to an increase in IL-10 mRNA expression in ALI, suggesting a beneficial combined action in modulating the inflammatory response.
Considering the correlation with humans, it is important to observe the effects in our study by performing training in the target zone of 70% of maximum VO2 or maximum heart rate. However, for humans, this training zone is individual (due to factors such as fitness level, age, and comorbidities) and should be determined through a treadmill exercise test (Fletcher et al. 2001) or using formulas based on the Karvonen model (She et al. 2015). Therefore, the treadmill speed varies from person to person.
Although we observed a decreased in ALI in male mice, when pre-treated with L-thyroxine and Ex, the same should not be directly assumed for female mice as they could present different inflammatory responses. Our study was conducted with only male mice to minimize hormonal variability, since males exhibit more stable hormone levels compared to females, who experience hormonal fluctuations due to the estrous cycle, and to reduce initial behavioral and physiological variability (Scotland et al. 2011; Villar et al. 2011; Bojalil et al. 2023). Additionally, inflammatory response methodologies usually employ male mice (Gholamnezhad et al. 2022). We recognize that our approach limits the generalization of our findings to both sexes, and therefore our results serve as a basis for future studies that will include both males and females.
Conclusions
In conclusion, our results provide evidence that L-thyroxine has an immunomodulatory capacity in LPS-stimulated macrophages and this effect is maintained in association with Ex in an LPS-induced ALI model in mice. Pre-treatments with L-thyroxine, as well as its combination with Ex, resulted in reduced lung injury through cell displacement and expression of inflammatory cytokines. It is crucial to emphasize that the combination of pre-treatments with L-thyroxine and Exercise did not potentiate the anti-inflammatory effect of L-thyroxine. The search for preventive and therapeutic strategies for ALI is of paramount importance since, once the disease is installed, treatment options are mainly restricted to supportive measures. Therapies that aim to reduce inflammation in the lungs and prevent disease progression can be particularly helpful in combating ALI. We have shown that the pre-treatment with L-thyroxine, as well as its association with Ex, can contribute to the protection of lung cells. Further research is needed to gain a more complete understanding of the mechanism by which L-thyroxine regulates the inflammatory response during ALI. In addition, this study contributes to the knowledge of the anti-inflammatory activities of L-thyroxine, expanding research perspectives in this area.
Acknowledgements
Coordenação de Aperfeiçoamento de Pessoal de Nível Superior—Brasil (CAPES) and Pontifícia Universidade Católica do Rio Grande do Sul (PUCRS).
References
Abbas A.M., Sakr H.F. 2016. Effect of magnesium sulfate and thyroxine on inflammatory markers in a rat model of hypothyroidism. Canadian Journal of Physiology and Pharmacology, 94(4): 426–432.
Abd El-Kader S., Gari A., Salah El-Den A. 2013. Impact of moderate versus mild aerobic exercise training on inflammatory cytokines in obese type 2 diabetic patients: a randomized clinical trial. African Health Sciences, 13(4): 857–863.
Abd El-Kader S.M., Al-Shreef F.M. 2018. Inflammatory cytokines and immune system modulation by aerobic versus resisted exercise training for elderly. African Health Sciences, 18(1): 120–131.
Al-Abed Y., Metz C.N., Cheng K.F., Aljabari B., VanPatten S., Blau S., et al. 2011. Thyroxine is a potential endogenous antagonist of macrophage migration inhibitory factor (MIF) activity. Proceedings of the National Academy of Sciences, 108(20): 8224–8227.
Aquim E.E., Bernardo W.M., Buzzini R.F., Azeredo N.S.G., Cunha L.S.D., Damasceno M.C.P., et al. 2019. Brazilian guidelines for early mobilization in Intensive care unit. Revista Brasileira de Terapia Intensiva, 31(4): 434–443.
Arnett D.K., Blumenthal R.S., Albert M.A., Buroker A.B., Goldberger Z.D., Hahn E.J., et al. 2019. 2019 ACC/AHA Guideline on the primary prevention of Cardiovascular Disease: A report of the American College of Cardiology/American Heart Association Task Force on clinical Practice Guidelines. Circulation, 140(11): e596–e646.
Baek S.H., Park T., Kang M.G., Park D. 2020. Anti-inflammatory activity and ROS regulation effect of sinapaldehyde in LPS-stimulated RAW 264.7 macrophages. Molecules (Basel, Switzerland), 25(18): 4089.
Bavarsad K., Hadjzadeh M.A., Hosseini M., Pakdel R., Beheshti F., Bafadam S., Ashaari Z. 2020. Effects of levothyroxine on learning and memory deficits in a rat model of Alzheimer's disease: the role of BDNF and oxidative stress. Drug and Chemical Toxicology, 43(1): 57–63.
Bojalil R., Ruíz-Hernández A., Villanueva-Arias A., Amezcua-Guerra L.M., Cásarez-Alvarado S., Hernández-Dueñas A.M., et al. 2023. Two murine models of sepsis: immunopathological differences between the sexes-possible role of TGFβ1 in female resistance to endotoxemia. Biological Research, 56(1): 54.
Booth F.W., Roberts C.K., Laye M.J. 2012. Lack of exercise is a major cause of chronic diseases. Comprehensive Physiology, 2(2): 1143–1211.
Boucher P.E., Taplin J., Clement F. 2022. The cost of ARDS: a systematic review. Chest, 161(3): 684–696.
Bruunsgaard H. 2005. Physical activity and modulation of systemic low-level inflammation. Journal of Leukocyte Biology 78(4): 819–835.
Chaker L., Bianco A.C., Jonklaas J., Peeters R.P. 2017. Hypothyroidism. The Lancet, 390(10101): 1550–1562.
Chakrabarti S.K., Ghosh S., Banerjee S., Mukherjee S., Chowdhury S. 2016. Oxidative stress in hypothyroid patients and the role of antioxidant supplementation. Indian Journal of Endocrinology and Metabolism, 20(5): 674–678.
Chen Y., Sjölinder M., Wang X., Altenbacher G., Hagner M., Berglund P., et al. 2012. Thyroid hormone enhances nitric oxide-mediated bacterial clearance and promotes survival after meningococcal infection. PLoS ONE, 7(7): e41445.
Cheserek M.J., Wu G.R., Ntazinda A., Shi Y.H., Shen L.Y., Le G.W. 2015. Association between thyroid hormones, lipids and oxidative stress markers in subclinical hypothyroidism. Journal of Medical Biochemistry, 34(3): 323–331.
De Vito P., Balducci V., Leone S., Percario Z., Mangino G., Davis P.J., et al. 2012. Nongenomic effects of thyroid hormones on the immune system cells: new targets, old players. Steroids, 77(10): 988–995.
Fan E., Dowdy D.W., Colantuoni E., Mendez-Tellez P.A., Sevransky J.E., Shanholtz C., et al. 2014. Physical complications in acute lung injury survivors: a two-year longitudinal prospective study. Critical Care Medicine, 42(4): 849–859.
Fei J., Fu L., Hu B., Chen Y.H., Zhao H., Xu D.X., Li J.B. 2019. Obeticholic acid alleviate lipopolysaccharide-induced acute lung injury via its anti-inflammatory effects in mice. International Immunopharmacology, 66: 177–184.
Fletcher G.F., Balady G.J., Amsterdam E.A., Chaitman B., Eckel R., Fleg J., et al. 2001. Exercise standards for testing and training: a statement for healthcare professionals from the American Heart Association. Circulation, 104(14): 1694–1740.
Furuya F., I. T., Tamura S., Takahashi K., Kobayashi H., et al. 2017. The ligand-bound thyroid hormone receptor in macrophages ameliorates kidney injury via inhibition of nuclear factor-κb activities. 7.
Gajic O., Dabbagh O., Park P.K., Adesanya A., Chang S.Y., Hou P., et al. 2011. Early identification of patients at risk of acute lung injury: evaluation of lung injury prediction score in a multicenter cohort study. American Journal of Respiratory and Critical Care Medicine, 183(4): 462–470.
Gholamnezhad Z., Safarian B., Esparham A., Mirzaei M., Esmaeilzadeh M., Boskabady M.H. 2022. The modulatory effects of exercise on lipopolysaccharide-induced lung inflammation and injury: a systemic review. Life Sciences, 293: 120306.
Gleeson M., Bishop N.C., Stensel D.J., Lindley M.R., Mastana S.S., Nimmo M.A. 2011. The anti-inflammatory effects of exercise: mechanisms and implications for the prevention and treatment of disease. Nature Reviews Immunology, 11(9): 607–615.
Grommes J., Soehnlein O. 2011. Contribution of neutrophils to acute lung injury. Molecular Medicine, 17(3-4): 293–307.
Haute G.V., Luft C., Pedrazza L., Antunes G.L., Silveira J., de Souza Basso B., et al. 2023. Simvastatin attenuates inflammatory process on LPS-induced acute lung injury in mice. Respiratory Physiology and Neurobiology, 309: 104002.
Huang M., Parker A.M., Bienvenu O.J., Dinglas V.D., Colantuoni E., Hopkins R.O., et al. 2016. Psychiatric symptoms in acute Respiratory distress syndrome survivors: A 1-year national multicenter study. Critical Care Medicine, 44(5): 954–965.
Improta-Caria A.C., Soci Ú., Pinho C.S., Aras Júnior R., De Sousa R.A.L., Bessa T.C.B. 2021. Physical exercise and immune system: perspectives on the COVID-19 pandemic. Revista da Associação Médica Brasileira, (1992) 67(Suppl 1): 102–107.
Jin S.C., Kim M.H., Jo S.Y., Yoon Choi L., Lee H., Yang W.M. 2020. Soshiho-tang protects LPS-induced acute liver injury by attenuating inflammatory response. Journal of Natural Medicines, 74(4): 788–795.
Kalyanaraman B., Darley-Usmar V., Davies K.J., Dennery P.A., Forman H.J., Grisham M.B., et al. 2012. Measuring reactive oxygen and nitrogen species with fluorescent probes: challenges and limitations. Free Radical Biology and Medicine, 52(1): 1–6.
Kedare S.B., Singh R.P. 2011. Genesis and development of DPPH method of antioxidant assay. Journal of Food Science and Technology, 48(4): 412–422.
Knaus W.A., Draper E.A., Wagner D.P., Zimmerman J.E. 1985. APACHE II: a severity of disease classification system. Critical Care Medicine, 13(10): 818–829.
Lang X., Zhao N., He Q., Li X., Sun C., Zhang X. 2020. Treadmill exercise mitigates neuroinflammation and increases BDNF via activation of SIRT1 signaling in a mouse model of T2DM. Brain Research Bulletin, 165: 30–39.
Lin S., Wu H., Wang C., Xiao Z., Xu F. 2018. Regulatory T cells and acute lung injury: cytokines, uncontrolled inflammation, and therapeutic implications. Frontiers in Immunology, 9: 1545.
Liu T., Zhang L., Joo D., Sun S.C. 2017. NF-κb signaling in inflammation. Signal Transduction and Targeted Therapy, 2: 17023–.
Livak K.J., Schmittgen T.D. 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods (San Diego, Calif.) 25(4): 402–408.
Magri F., Buonocore M., Oliviero A., Rotondi M., Gatti A., Accornero S., et al. 2010. Intraepidermal nerve fiber density reduction as a marker of preclinical asymptomatic small-fiber sensory neuropathy in hypothyroid patients. European Journal of Endocrinology, 163(2): 279–284.
Marfella R., Ferraraccio F., Rizzo M.R., Portoghese M., Barbieri M., Basilio C., et al. 2011. Innate immune activity in plaque of patients with untreated and L-thyroxine-treated subclinical hypothyroidism. The Journal of Clinical Endocrinology and Metabolism, 96(4): 1015–1020.
Martha B.A., Vacchi C.O., Fattori R.A., Macagnan F.E. 2021. Effect of physical exercise on the functional capacity of children and adolescents submitted to transplantation of hematopoietic stem cells-A systematic review with meta-analysis. Journal of Child Health Care, 25(1): 18–30.
McAninch E.A., Bianco A.C. 2014. Thyroid hormone signaling in energy homeostasis and energy metabolism. Annals of the New York Academy of Sciences, 1311: 77–87.
Meduri G.U., Kohler G., Headley S., Tolley E., Stentz F., Postlethwaite A. 1995. Inflammatory cytokines in the BAL of patients with ARDS. Persistent elevation over time predicts poor outcome. Chest, 108(5): 1303–1314.
Mowery N.T., Terzian W.T.H., Nelson A.C. 2020. Acute lung injury. Current Problems in Surgery, 57(5): 100777.
Mullur R., Liu Y.Y., Brent G.A. 2014. Thyroid hormone regulation of metabolism. Physiological Reviews, 94(2): 355–382.
Ning Y., Zhang M., DU Y.H., Zhang H.N., Li L.Y., Qin Y.W., et al. 2018. [Effects of thyroid hormone on macrophage dysfunction induced by oxidized low-density lipoprotein]. Sheng Li Xue Bao [Acta Physiologica Sinica], 70(2): 141–148.
Pierce A., Pittet J.F. 2014. Inflammatory response to trauma: implications for coagulation and resuscitation. Current Opinion in Anaesthesiology, 27(2): 246–252.
Qin X., Jiang X., Wang Y., Miao Z., He W., Yang G., et al. 2016. Micheliolide inhibits LPS-induced inflammatory response and protects mice from LPS challenge. Scientific Reports, 6: 23240.
Ranieri V.M., Rubenfeld G.D., Thompson B.T., Ferguson N.D., Caldwell E., Fan E., et al. 2012. Acute respiratory distress syndrome: the Berlin definition. Jama, 307(23): 2526–2533.
Robb C.T., Regan K.H., Dorward D.A., Rossi A.G. 2016. Key mechanisms governing resolution of lung inflammation. Seminars in Immunopathology, 38(4): 425–448.
Saguil A., Fargo M.V. 2020. Acute respiratory distress syndrome: diagnosis and management. American Family Physician, 101(12): 730–738.
Santi A., Duarte M.M., Moresco R.N., Menezes C., Bagatini M.D., Schetinger M.R., Loro V.L. 2010. Association between thyroid hormones, lipids and oxidative stress biomarkers in overt hypothyroidism. cclm 48(11): 1635–1639.
Scheffer D.D.L., Ghisoni K., Aguiar A.S., Latini A. 2019. Moderate running exercise prevents excessive immune system activation. Physiology and Behavior, 204: 248–255.
Scotland R.S., Stables M.J., Madalli S., Watson P., Gilroy D.W. 2011. Sex differences in resident immune cell phenotype underlie more efficient acute inflammatory responses in female mice. Blood, 118(22): 5918–5927.
Seyedhosseini Tamijani S.M., Beirami E., Ahmadiani A., Dargahi L. 2019. Thyroid hormone treatment alleviates the impairments of neurogenesis, mitochondrial biogenesis and memory performance induced by methamphetamine. Neurotoxicology, 74: 7–18.
She J., Nakamura H., Makino K., Ohyama Y., Hashimoto H. 2015. Selection of suitable maximum-heart-rate formulas for use with Karvonen formula to calculate exercise intensity. International Journal of Automation and Computing, 12: 62–69.
Sáez de Asteasu M.L., Martínez-Velilla N., Zambom-Ferraresi F., Casas-Herrero Á., Lucía A., Galbete A., Izquierdo M. 2019. Physical exercise improves function in acutely hospitalized older patients: secondary analysis of a randomized clinical trial. Journal of the American Medical Directors Association, 20(7): 866–873.
Tang J., Diao P., Shu X., Li L., Xiong L. 2019. Quercetin and quercitrin attenuates the inflammatory response and oxidative stress in LPS-induced RAW264.7 cells: In vitro assessment and a theoretical model. BioMed Research International, 2019: 1.
Tayde P.S., Bhagwat N.M., Sharma P., Sharma B., Dalwadi P.P., Sonawane A., et al. 2017. Hypothyroidism and depression: are cytokines the link? Indian Journal of Endocrinology and Metabolism, 21(6): 886–892.
Tellechea M.L. 2021. Meta-analytic evidence for increased low-grade systemic inflammation and oxidative stress in hypothyroid patients. Can levothyroxine replacement therapy mitigate the burden? Endocrine, 72(1): 62–71.
Tu H., Zhang D., Barksdale A.N., Wadman M.C., Muelleman R.L., Li Y.L. 2020. Dexamethasone improves wound healing by decreased inflammation and increased vasculogenesis in mouse skin frostbite model. Wilderness and Environmental Medicine, 31(4): 407–417.
Villar I.C., Scotland R.S., Khambata R.S., Chan M., Duchene J., Sampaio A.L., et al. 2011. Suppression of endothelial P-selectin expression contributes to reduced cell trafficking in females: an effect independent of NO and prostacyclin. Arteriosclerosis, Thrombosis, and Vascular Biology, 31(5): 1075–1083.
Vincent J.L., Moreno R., Takala J., Willatts S., De Mendonça A., Bruining H., et al. 1996. The SOFA (Sepsis-related Organ Failure Assessment) score to describe organ dysfunction/failure. On behalf of the Working Group on Sepsis-Related Problems of the European Society of Intensive Care Medicine. Intensive Care Medicine, 22(7): 707–710.
Voiriot G., Razazi K., Amsellem V., Tran Van Nhieu J., Abid S., Adnot S., et al. 2017. Interleukin-6 displays lung anti-inflammatory properties and exerts protective hemodynamic effects in a double-hit murine acute lung injury. Respiratory Research, 18(1): 64.
Wang X., Wang Z., Tang D. 2021. Aerobic exercise improves LPS-induced sepsis via regulating the Warburg effect in mice. Scientific Reports, 11(1): 17772.
Wenzek C., Boelen A., Westendorf A.M., Engel D.R., Moeller L.C., Führer D. 2022. The interplay of thyroid hormones and the immune system—where we stand and why we need to know about it. European Journal of Endocrinology, 186(5): R65–R77.
Werneck F.Z., Coelho E.F., Almas S.P., Garcia M.M.D.N., Bonfante H.L.M., Lima J.R.P., et al. 2018. Exercise training improves quality of life in women with subclinical hypothyroidism: a randomized clinical trial. Archives of Endocrinology and Metabolism, 62(5): 530–536.
Winters B.D., Eberlein M., Leung J., Needham D.M., Pronovost P.J., Sevransky J.E. 2010. Long-term mortality and quality of life in sepsis: a systematic review. Critical Care Medicine, 38(5): 1276–1283.
Wu C.W., Chen Y.C., Yu L., Chen H.I., Jen C.J., Huang A.M., et al. 2007. Treadmill exercise counteracts the suppressive effects of peripheral lipopolysaccharide on hippocampal neurogenesis and learning and memory. Journal of Neurochemistry, 103(6): 2471–2481.
Yi L., Zhou Z., Zheng Y., Chang M., Huang X., Guo F., et al. 2019. Suppressive effects of GSS on lipopolysaccharide-induced endothelial cell injury and ALI via TNF-. Mediators of Inflammation, 2019: 1.
Zarghi A., Arfaei S. 2011. Selective COX-2 inhibitors: a review of their structure-activity relationships. Iranian Journal of Pharmaceutical Research, 10(4): 655–683.
Zhang Z., Luo Z., Bi A., Yang W., An W., Dong X., et al. 2017. Compound edaravone alleviates lipopolysaccharide (LPS)-induced acute lung injury in mice. European Journal of Pharmacology, 811: 1–11.
Information & Authors
Information
Published In

FACETS
Volume 10 • Number 1 • January 2025
Pages: 1 - 14
Editor: Charles Couillard
History
Received: 29 December 2023
Accepted: 10 October 2024
Version of record online: 28 January 2025
Copyright
© 2025 The Authors. This work is licensed under a Creative Commons Attribution 4.0 International License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Data Availability Statement
Data and material will be provided by corresponding author if requested.
Key Words
Sections
Subjects
Plain Language Summary
L-thyroxine and its association with physical exercise protects against acute lung injury
Authors
Author Contributions
Conceptualization: BAM
Formal analysis: BAM, CL, BPC, GLA
Investigation: BAM, GVT, LSM, MCRG, JRdO
Methodology: BAM, GVT, CL, BPC, GLA, MSdC, LSM, MCRG
Resources: GVT, JRdO
Software: CL
Supervision: BAM
Visualization: BPC, MSdC, JRdO
Writing – original draft: BAM, JRdO
Writing – review & editing: BAM, JRdO
Competing Interests
The authors declare that they have no competing interests.
Funding Information
Metrics & Citations
Metrics
Other Metrics
Citations
Cite As
Bianca Andrade Martha, Giovana Vivan Tonial, Carolina Luft, Bruna Pasqualotto Costa, Géssica Luana Antunes, Mariana Severo da Costa, Lucas Strassburger Matzenbacher, Maria Cláudia Rosa Garcia, and Jarbas Rodrigues de Oliveira. 2025. The effects of L-thyroxine and its association with physical exercise decrease the inflammation in an acute lung injury model. FACETS.
10: 1-14.
https://doi.org/10.1139/facets-2023-0239
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
There are no citations for this item