Multifacets in silico study of Plant Defensin 1.1 protein (PDF1.1) in Brassica rapa (turnip)
Abstract
Brassica rapa (turnip) belongs to the Brassicaceae family. Due to the diseases caused by fungal pathogens such as Erysiphe cruciferarum, Alternaria japonica, and Alternaria brassicae, the yield of B. rapa was reduced in many Asian and European countries. Plant defensins (PDFs) are proteins that have antifungal properties. This research aimed to comprehend the key attributes of B. rapa PDF 1.1s by incorporating a wide range of in silico approaches. Initially, we explored orthologues of PDF 1.1 in B. rapa through Blastp in Phytozome v.13 which highlighted fourteen different orthologues. Afterward, by using Mega X, the homology of B. rapa PDF1.1 proteins with Arabidopsis thaliana PDF1.1 proteins was also inferred. While, the results of NCBI CD show that these proteins possessed a common domain, i.e., gamma-thionin. The major findings of this study included the crucial characteristics of PDF1.1 proteins that were explored by utilizing the PANZZER2 web server and BlastKOALA, which proposed that these proteins possessed degradation ability of pathogen’s cells and were also found to be involved in a mitogen-activated protein kinase signaling pathway. Overall, this research provided a better understanding of the crucial contribution of B. rapa PDFs 1.1 under biotic stress specifically, the pathogenic fungal attack.
1. Introduction
Brassica rapa (turnip) is an essential root and leaf crop cultivated worldwide and it belongs to the family Brassicaceae which approximately consists of 35 000 species. Delicious roots and green leaves are the reasons for the cultivation of B. rapa, its roots are used in making salads, and green leaves are consumed after cooking or steaming which are more beneficial for human health in comparison with turnip roots because the roots contain the least amount of phenolics (Dejanovic et al. 2021). In addition, B. rapa has some traditional beneficial effects like turnips are therapeutic agents for various types of liver and kidney diseases. Turnips have also pharmacological importance, they are antimicrobial, anti-inflammatory, antioxidant, antidiabetic, cardioprotective, nephroprotective, antitumor, and analgesic (Paul et al. 2019). Studies show that turnips are rich in glucosinates, sulfur compounds, carbohydrates, e.g., polysaccharides, and minerals, e.g., calcium 4500 mg per kg, zinc 15.7 mg per kg, and iron 81.1 mg per kg (). They also contain vitamin C, flavonoids, isothiocyanates, phenolics, volatiles, and saponins, and the slices of turnip contain 61%–39% of essential and nonessential amino acids (Sun et al. 2022).
Various pathogenic fungi drastically affect the growth and yield of different species of this plant. Some of Brassica species’ common and most impacting fungal diseases are powdery mildew, downy mildew, Alternaria leaf spot, white rust, and blackleg. The combination of Alternaria brassicae, Alternaria japonica, and Alternaria brassicicola causes the Black spot disease in B. rapa which decreases the germination and causes shuttering of premature pods, Albugo candida causes the white rust disease in B. rapa to which 60% of the yield was lost in Canada. Powdery mildew is caused by the pathogen Erysiphe cruciferarum in Brassica crops and lost yield worldwide (USA, Australia, and European and Asian countries) (Mourou et al. 2023).
Plants contain specified proteins designated as plant defensins (PDFs) to tackle such infections. These proteins are expressed in the cell wall of plant cells or extracellular spaces and are found in different parts of the plant such as seeds, leaves, flowers, roots, and stems (Yao et al. 2019). PDFs are small cysteine-rich proteins mainly consisting of a cystine-rich domain, a variable region on the C-terminal, and a signal peptide on the N-terminal (Yao et al. 2019). In Arabidopsis thaliana, two families of PDFs were identified (PDF1 and PDF2). Several studies reported the organ-specific expression of PDF1.1 (Yao et al. 2019). PDFs are also involved in the detoxification of heavy metals. They were entitled to this name due to their function and structural homology to that of insect and mammal defensins. These proteins are triggered by the pathogenic attack or any sort of abiotic stress, most commonly the antifungal infection (Kovaleva et al. 2020). Different studies have reported their anti-fungal response in different plants. For example, one of the investigations has reported that PDFs of Nicotiana alata, Solanum lycopersicum, and Pisum sativum induced fungal cell death by disrupting the plasma membrane which ultimately inhibited the cell division (Sathoff et al. 2019).
In this particular study, we have illustrated various aspects of PDF1.1 protein in B. rapa using the Bioinformatics tools, to comprehend their major characteristics. In this context, phylogenetic analysis, analysis of a conserved domain, motifs, physicochemical characterization, subcellular localization, prediction of secondary structure, and homology modeling along with exploring the pockets in respective structures were predicted. The main objective was to find the gene ontology or biological and molecular function of PDF1.1 protein in B. rapa. This study’s key findings indicate that the PDF1.1 protein in B. rapa exhibits strong antifungal activity, potentially playing a role in activating the MAPK signaling pathway to enhance the plant’s defense response against fungal pathogens. By understanding the role of PDF1.1 in plant immunity, researchers may develop strategies to induce the expression of these genes in B. rapa, providing an effective means to bolster resistance. This work offers a foundation for future efforts in breeding or engineering Brassica varieties with enhanced pathogen resilience, which could lead to more sustainable crop protection and improved yields.
2. Materials and methods
2.1. Sequence retrieval and orthologue identification
The amino-acid sequences of PDF1.1 protein were retrieved from the Ensembl Plant Database (https://plants.ensembl.org/index.html) (Cao et al. 2020) That contains the genomic sequences of various plant species. Blastp was done by using a genomic portal where we analyze the evolutionary history at sequence, structure, and gene family level, Phytozome v.13 (https://phytozome-next.jgi.doe.gov/blast-search) (Li et al. 2023) used to find orthologues in B. rapa FPsc v1.3, Prunus persica v2.1, Triticum aestivum v2.2, and in Arabidopsis thaliana TAIR10. The specific transcript, genomic sequence, and CDS files of PDF1.1 from B. rapa were downloaded. The genomic IDs of B. rapa PDF genes are given in Table 1. Named the B. rapa PDF1.1 proteins as BRa.PDF1.1 for convenience.
Table 1.
| Gene identifier | Protein name | Genomic position | Strand | Protein length (a.a) | Molecular weight (kDa) | Isoelectric point (pI) | No. of negative amino acids | No. of positive amino acid | Instability index | Aliphatic index | GRAVY |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Brara.G02203 | BRa.PDF1.1a | A07: 19219502. .19219850 | – | 80 | 8820.41 | 8.70 | 4 | 8 | 29.49 | 78.12 | 0.294 |
| Brara.B02209 | BRa.PDF1.1b | A02: .14132354 .14132706 | – | 80 | 8684.22 | 8.47 | 4 | 7 | 32.96 | 78.25 | 0.280 |
| Brara.G03383 | BRa.PDF1.1c | A07: 25653708. .25654345 | + | 80 | 8830.37 | 8.70 | 3 | 7 | 29.45 | 78.12 | 0.336 |
| Brara.G02204 | BRa.PDF1.1d | A07: 19222899. .19223262 | – | 79 | 8623.13 | 8.48 | 4 | 7 | 27.83 | 77.85 | 0.386 |
| Brara.G03384 | BRa.PDF1.1e | A07: 25660401. .25660984 | + | 79 | 8567.07 | 8.15 | 4 | 6 | 32.86 | 81.65 | 0.477 |
| Brara.G03382 | BRa.PDF1.1f | A07: 25646658. .25647213 | + | 80 | 8956.56 | 8.47 | 3 | 6 | 14.82 | 87.88 | 0.349 |
| Brara.B02211 | BRa.PDF1.1 g | A02: 14142388. .14142994 | – | 80 | 8918.54 | 9.00 | 6 | 12 | 32.13 | 71.00 | 0.065 |
| Brara.B02210 | BRa.PDF1.1 h | A02: 14139327. .14139661 | – | 82 | 9240.08 | 9.24 | 3 | 11 | 43.41 | 73.90 | 0.237 |
| Brara.H02365 | BRa.PDF1.1i | A08: 20133196. .20133758 | + | 77 | 8757.23 | 8.85 | 4 | 9 | 51.39 | 59.61 | 0.082 |
| Brara.F03084 | BRa.PDF1.1j | A06: 24591867. .24592198 | – | 80 | 9288.20 | 9.41 | 6 | 15 | 42.69 | 70.75 | 0.033 |
| Brara.E02038 | BRa.PDF1.1k | A05: 18660288. .18660931 | – | 79 | 8653.89 | 5.45 | 7 | 5 | 38.33 | 76.58 | 0.272 |
| Brara.G03385 | BRa.PDF1.1l | A07: 25662643. .25663252 | + | 57 | 6162.26 | 5.05 | 6 | 4 | 35.31 | 99.47 | 0.721 |
| Brara.B02477 | BRa.PDF1.1 m | A02: 16886467. .16887149 | + | 75 | 8678.74 | 9.57 | 3 | 13 | 21.81 | 89.60 | 0.236 |
| Brara.G00903 | BRa.PDF1.1n | A07: 10770494. .10771091 | – | 130 | 13867.87 | 5.12 | 11 | 7 | 49.79 | 57.00 | −0.060 |
Table 2.
| Protein names | Phosphorylation sites | N-glycosylation Sites |
|---|---|---|
| BRa.PDF1.1a | 2 | 0 |
| BRa.PDF1.1b | 5 | 0 |
| BRa.PDF1.1c | 5 | 0 |
| BRa.PDF1.1d | 7 | 0 |
| BRa.PDF1.1e | 5 | 0 |
| BRa.PDF1.1f | 6 | 0 |
| BRa.PDF1.1 g | 5 | 0 |
| BRa.PDF1.1 h | 6 | 0 |
| BRa.PDF1.1i | 5 | 0 |
| BRa.PDF1.1j | 15 | 0 |
| BRa.PDF1.1k | 3 | 0 |
| BRa.PDF1.1l | 7 | 0 |
| BRa.PDF1.1 m | 5 | 0 |
| BRa.PDF1.1n | 6 | 0 |
2.2. Phylogenetic analysis
To analyze the phylogenetic relationship between the PDF1.1 proteins of B. rapa FPsc v1.3, Prunus persica v2.1, Triticum aestivum v2.2, and Arabidopsis thaliana TAIR10, the Mega-X software was used where we upload the FASTA file that carries the amino-acid sequences of all these proteins and aligned by selecting option, i.e., multiple sequence alignment (Stecher et al. 2020). By using the results of alignment, a phylogenetic tree was created with Maximum likelihood with 300 Bootstrap values, and the Jones–Taylor–Thornton (JTT) model. The radial tree was constructed to view the clades separately in different colors using the itol (https://itol.embl.de/) (Letunic and Bork 2021).
2.3. Physicochemical properties and structure prediction
The physicochemical properties of BRa.PDF1.1 proteins, including the amino acid chain length, molecular weight (kDa), number of negatively and positively charged residues, isoelectric point (pI), instability index, GRAVY, and aliphatic index, are predicted by using the ExPASy-ProtParam tool (https://web.expasy.org/protparam/) (Garg et al. 2016). This particular tool takes the amino-acid sequence of the protein to be analyzed in FASTA format and then processes it accordingly. However, the results are presented in the form of different percentages and scores in tabular form.
2.4. Analysis of subcellular localization
To predict the location of BRa.PDF1.1 proteins within the cell, WoLF PSORT (https://wolfpsort.hgc.jp/) (Jiang et al. 2021) were used and the heatmap that contains the combined results was generated by applying the TBtool (https://github.com/CJ-Chen/TBtools) (Chen and Xia 2022).
2.5. Chromosomal mapping
The chromosomal mapping of BRa.PDF1.1 genes in B. rapa were predicated by using the Mapgene2chromosome (MG2C) web v2.1 database (http://mg2c.iask.in/mg2c_v2.0/) (Chao et al. 2021). This particular tool enables us to analyze that at which location and at which arm of chromosome the required gene would be located.
2.6. Protein domain, motif, and gene structure prediction
NCBI Conserved Domain database (https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi) (Yang et al. 2020) was used to predict the protein family and conserved domain of the BRa.PDF1.1 protein which are the conserved sequences associated with the specific function. To find motifs that are the small sequence of amino acids in the BRa.PDF1.1 protein sequences, MEME suite version 5.5.1 (https://meme-suite.org/meme/) (Nystrom and McKay 2021) was used by selecting the maximum 20 motifs limit that plays an essential role in protein structure and function. However, the gene structure display server 2.0 (http://gsds.gao-lab.org/) (Hu et al. 2015) was used to generate the intron/exon schematic diagram of BRa.PDF1.1 genes or to determine the exact number of coding and noncoding regions within the gene.
2.7. Gene ontology (GO) and KEGG analysis
The PANZZER2 web server (http://ekhidna2.biocenter.helsinki.fi/sanspanz/) (Törönen and Holm 2022) was used to predict the gene ontology or functions of the BRa.PDF1.1 protein at biological, molecular, and cellular levels with positive predicted values (PPVs). The KEGG analysis was done by using the BlastKOALA web server (https://www.kegg.jp/blastkoala/) (Kanehisa and Sato 2020). By this analysis, we can predict and visualize the cascade like how the protein is activated in which stress and involved in which pathway.
2.8. Prediction of functional sites in PDF1.1 proteins
The exploration of functional sites that include phosphorylation and glycosylation sites in PDF1.1 proteins (Table 2) was achieved by using the NetPhos 3.1 server (https://services.healthtech.dtu.dk/services/NetPhos-3.1/) (Li et al. 2018), selecting the score >0.5, which shows the phosphorylation with related kinases and NetNGlyc 1.0 server (https://services.healthtech.dtu.dk/services/NetNGlyc-1.0/) (Joshi and Gupta 2015), respectively.
2.9. Secondary structure prediction, homology modeling, model evaluation, and identification of active sites
The SOPMA online server (https://npsa.lyon.inserm.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_sopma.html) (Angamuthu and Piramanayagam 2017) was used for the comprehensive secondary structure analysis of BRa.PDF1.1 protein which includes alpha helices, beta turns, beta strands, random coils, etc. This server also processes the FASTA format amino-acid sequence of the given protein and presents the results in different forms (peaks and bars). However, for homology modeling or three-dimensional (3D) structure prediction the BRa.PDF1.1 proteins, the Phyre (Paul et al. 2019) web server (http://www.sbg.bio.ic.ac.uk/phyre2) was used. After the prediction of 3D model of a protein, it is always necessary to validate its quality. For this purpose, Ramachandran plot was employed which is available on PROCHECK server (https://saves.mbi.ucla.edu/) (Laskowski et al. 2013). This tool analyzes the given protein on the basis of the stereochemical properties (analyzing the torsional angles phi and psi) and demonstrates the results in the form of a map, comprises of four different quadrants. Each quadrant provides the details regarding the most favorable, additionally allowed, generously allowed and unfavored regions, depending on the steric clashes, the respective amino acids of the given protein possess. However, the active protein pocket sites were predicted by using the CASTp tool (http://sts.bioe.uic.edu/castp/calculation.html) (Tian et al. 2018). This specific tool predicts different active sites or pockets of varied area and volume sizes that are presented in different colors. In context of a molecular interaction, these sites are responsible for interaction with other proteins or any other interacting molecules.
The following figure illustrates the whole workflow which is followed in this work.
3. Results
3.1. Phylogenetic analysis of PDF1.1 proteins
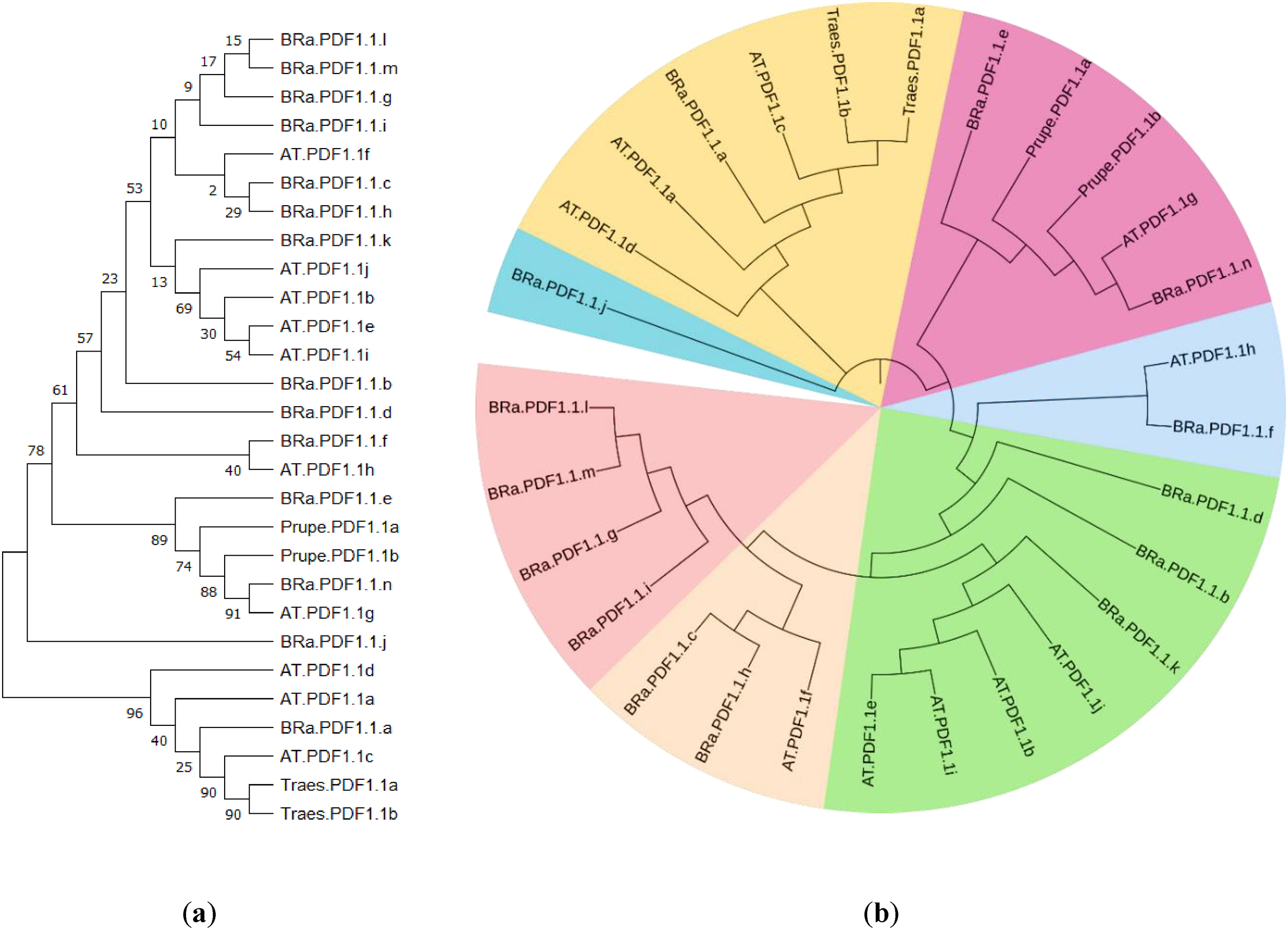
The phylogenetic analysis of PDF 1.1 proteins was constructed by employing the maximum-likelihood method which exhibited thought-provoking results. The selection of Prunus persica and Triticum aestivum as variants for constructing the phylogenetic tree (Fig. 1) was based on the orthology aspect of PDF1.1 proteins. These two species exhibited close orthology than the rest of the any other specie that is why P. persica and T. aestivum were specifically chosen. In this particular phylogenetic tree, a total of seven different clades were produced. This tree was further annotated to be converted into radial form to present in a bit more tangible manner. Each clade was presented with their respective colors to discriminate them from each other. This tree was constructed to infer the phylogenetic relationship of PDF1.1 proteins of B. rapa with those of Arabidopsis thaliana, Prunus persica, and Triticum aestivum. Clade 1 is designated with a light blue color, comprised of just two members, i.e., AT.PDF.1h and Bra.PDF1.1.f; Clade 2 was the largest among the rest of the others. It comprised various variants of PDF1.1 proteins of B. rapa and Arabidopsis thaliana. Clade 3 contained just one AT.PDF1.1f protein which exhibited its evolutionary association with PDF1.1.h and PDF1.1.c proteins of B. rapa. However, Clades 4 and 5 represented unique phylogeny behaviors. Clade 4 comprised four entities and all were different variants of Bra.PDF1.1 proteins. All of them were found in evolutionary association with each other. Clade 5 had only one entity i.e., BRaPDF1.1.j. The second largest clade in the phylogenetic tree was Clade 6, which is represented with a mild brown color in Fig. 2. This group displayed a diverse evolutionary linkage among PDF1.1 proteins of Arabidopsis thaliana, B. rapa, and Triticum aestivum. Clade 7 also showed a similar relatedness fashion of PDF1.1 proteins which belong to B. rapa, Arabidopsis thaliana, and Prunus persica. In conclusion, B. rapa PDF1.1 demonstrates greater similarity to Arabidopsis thaliana PDF1.1 than the PDF1.1 proteins of T. aestivum because multiple evolutionary linkages were detected at various clades of the phylogenetic tree.
Fig. 1.
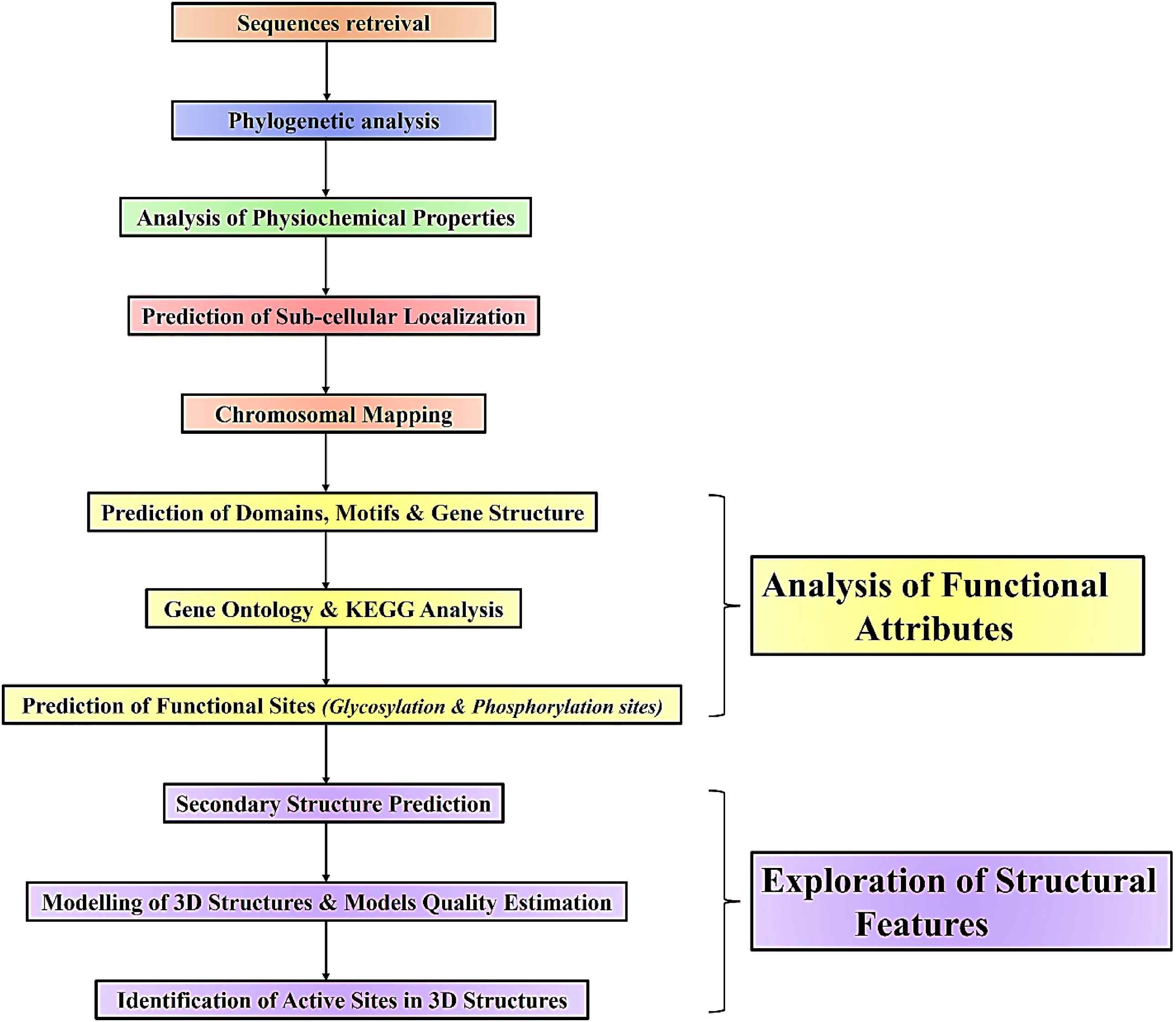
Fig. 2.
3.2. Analysis of physicochemical properties
The results of physicochemical characterization exhibited that the BRa.PDF1.1 proteins possessed mostly positively charged amino acids with protein lengths ranging between 57 and 130, and molecular weights varying from 6162.26 to 13867.87 kDa, while their instability index (II) ranges between 14.82 and 51.39. Furthermore, the BRa.PDF1.1 proteins GRAVY ranged from 0.060 to 0.72, their isoelectric point (pI) ranged from 5.05 to 9.57, and their aliphatic index ranged from 57.00 to 99.47. The compiled results of the physicochemical properties of all these proteins are summarized in Table 1.
3.3. Subcellular localization prediction of Bra.PDF1.1s
The subcellular analysis of the proteins under study was performed by WoLF PSORT. It presented the results in the form of a heat map. The different colors in the heat map indicated the relevant scores concerning the localities of these proteins in various cellular and extracellular compartments. These results suggested that the BRa.PDF1.1 proteins are primarily located in the extracellular spaces but some of them are also located in chloroplast, cytosol, endoplasmic reticulum, nucleus, vacuoles, and golgi bodies. The graphical demonstration of this analysis is illustrated in Fig. 3. The names of the proteins and the cellular compartments are mentioned along the X-axis and Y-axis, respectively. A colored scheme is also given which contains different confidence scores for correspondence to infer the proteins’ respective localization.
Fig. 3.
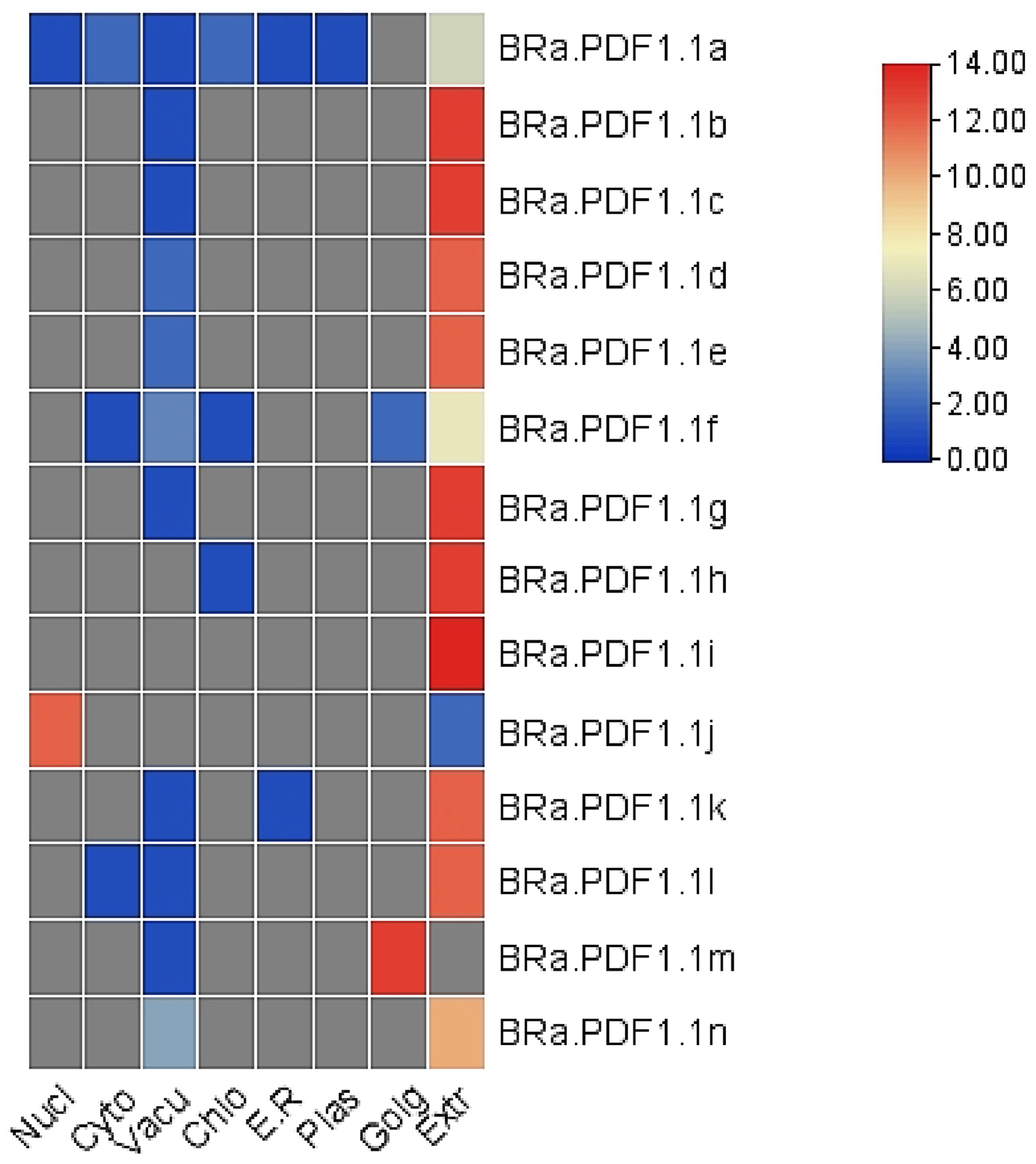
3.4. Chromosomal localization of Bra.PDF1.1 genes
The localization of B. rapa PDF1.1 genes on chromosomes was predicted by using the Mapgene2choromosome web v2.1 database and is represented in Fig. 4. The results of this chromosomal mapping showed that the maximum number of genes are located on chromosome no. 7. Four genes were found to be located on chromosome no. 2, one on chromosome no. 5, chromosome no. 6, and chromosome no. 8. However, seven genes are located on chromosome no. 7.
Fig. 4.
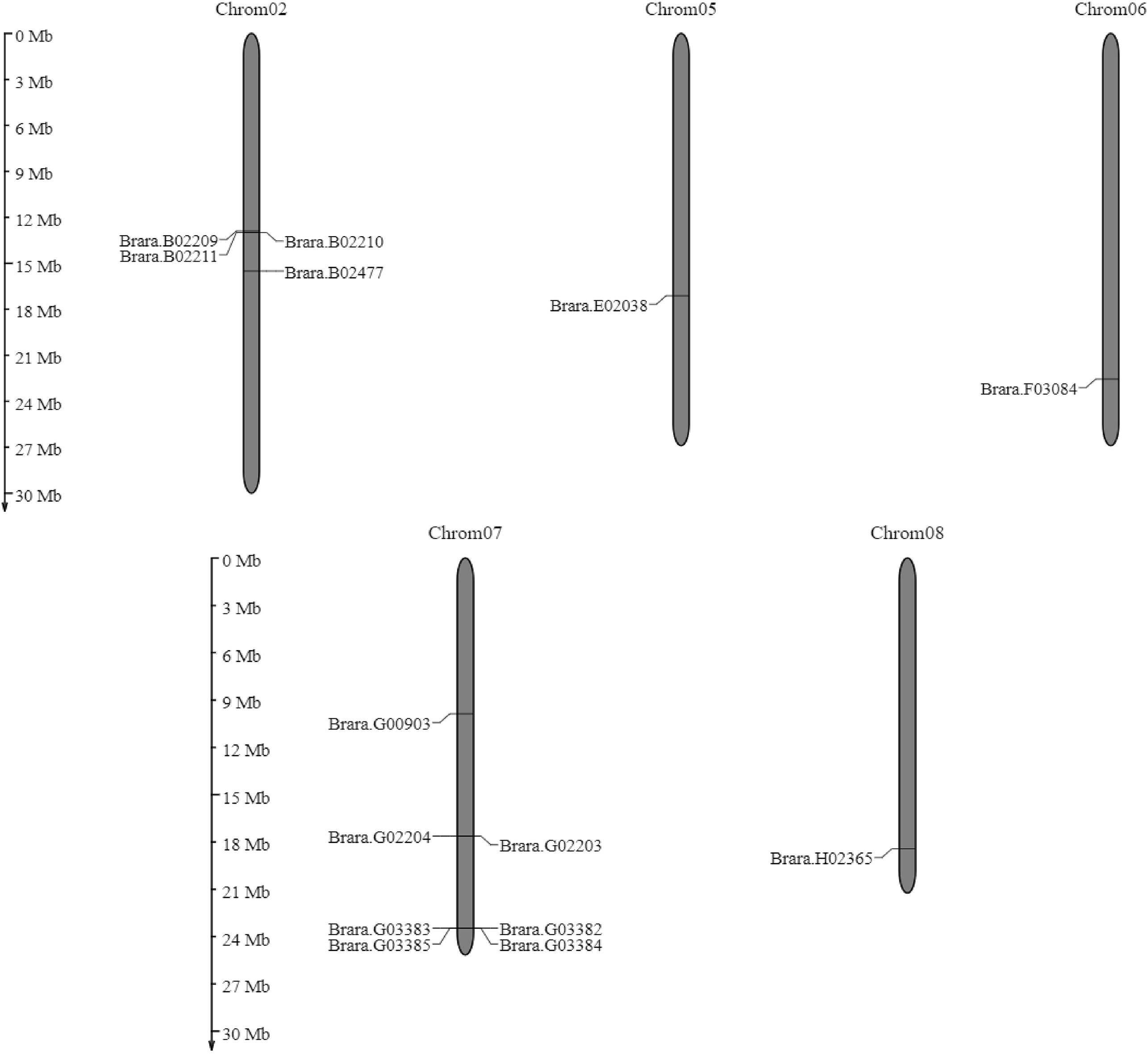
3.5. Prediction of protein domains, motifs, introns, and exons in Bra.PDF1.1s
Protein domains were identified by using the NCBI conserved domain database. Results showed that the BRa.PDF1.1 protein sequences possessed a common conserved domain, i.e., gamma thionin, and four of the query sequences showed no conserved domains. The function of the gamma-thionin family protein adopts the “knottin” fold, a stable cysteine-rich scaffold, in which one disulfide bridge crosses the macrocycle made by two other disulfide bridges and connects segments of the backbone. They are found in plant lectins/antimicrobial peptides, plant proteinase/amylase inhibitors, plant gamma-thionins, and arthropod defensins. Figure 5 represents the results of the domain analysis of BRa.PDF1.1s.
Fig. 5.
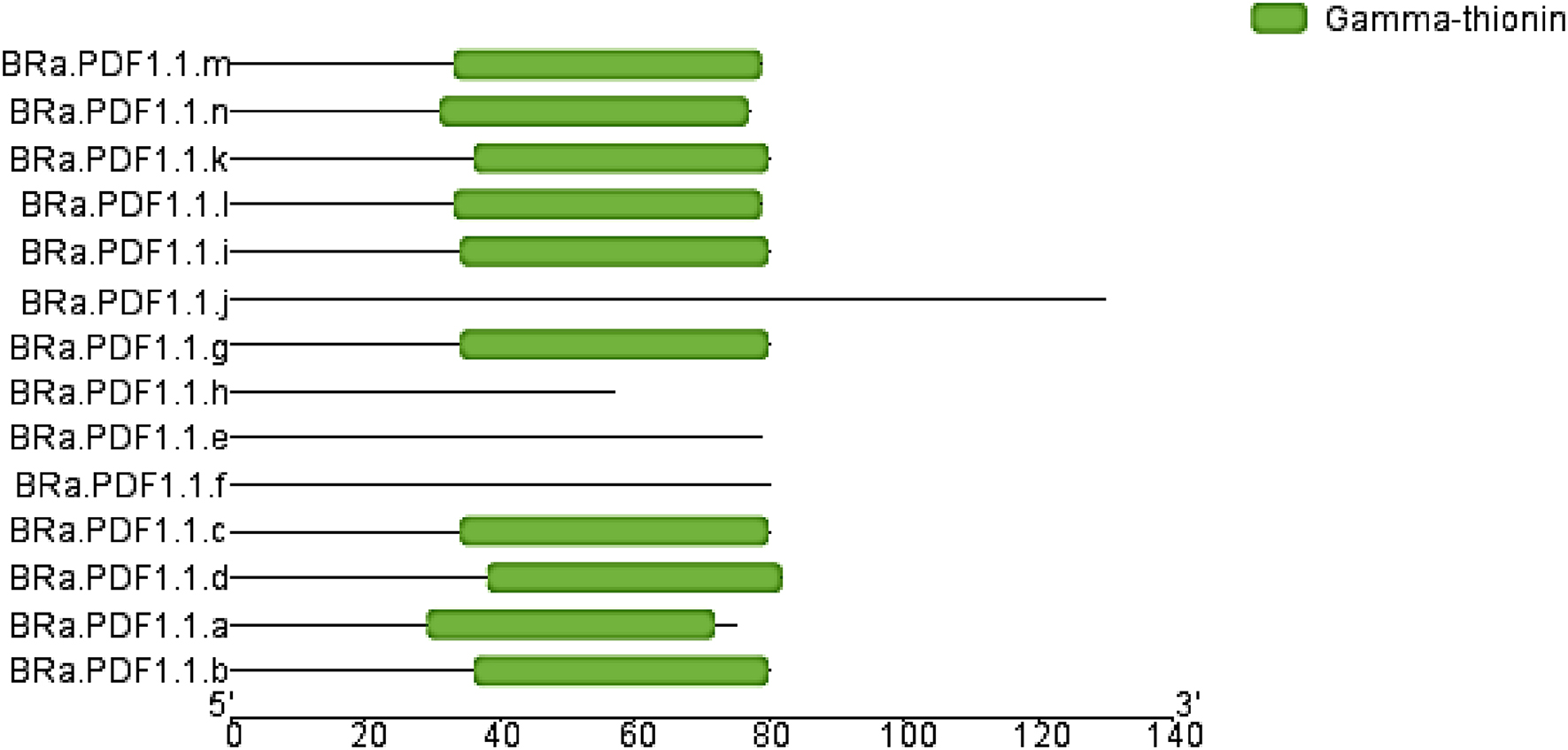
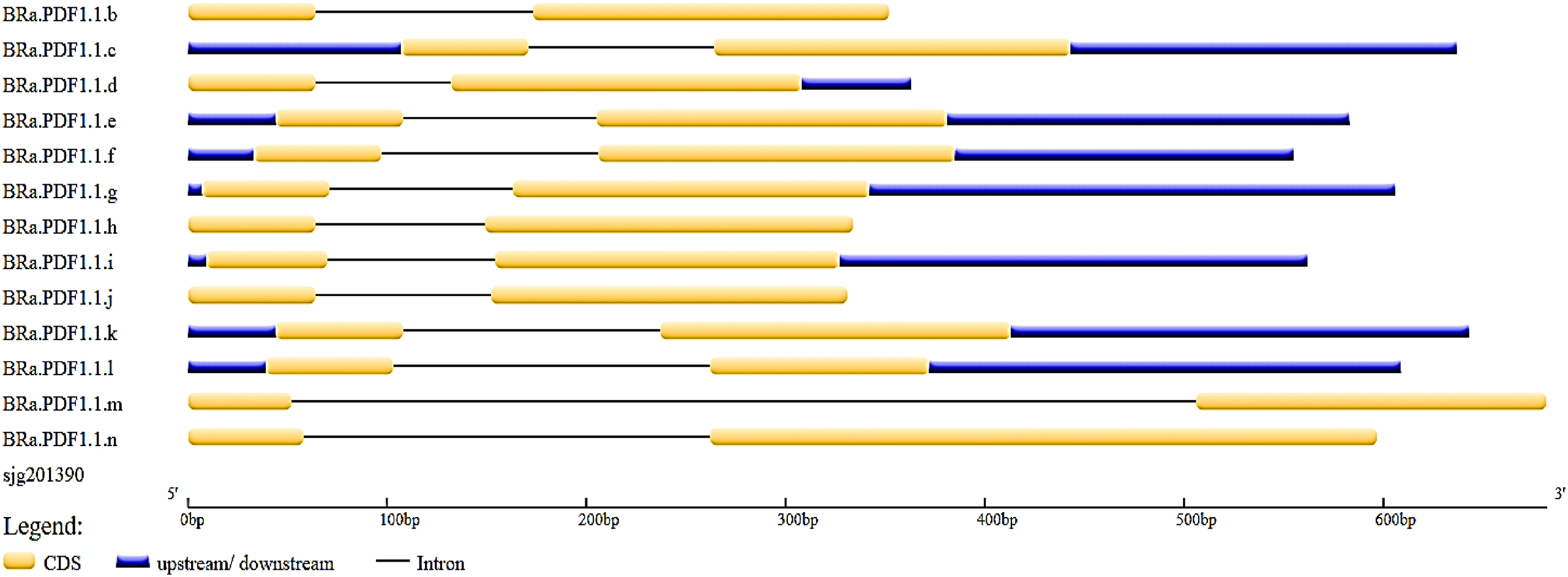
On the other hand, the protein motifs were identified by using the MEME suite version 5.5.1. The results of motifs prediction are presented in Fig. 4, which illustrates the motif locations along their corresponding p-values. Notably, 20 motifs were found in the amino-acid sequences of the proteins under study. Each motif displays a specific sequence and is represented with various colors which can be observed in Fig. 6 and Fig. 7. The intron/exon organization of BRa.PDF1.1 genes were performed by using the Gene Structure Display Server 2.0. All of the genes comprised two exons that are represented in yellow color and one intron which is represented in black lines as presented in Fig. 8.
Fig. 6.
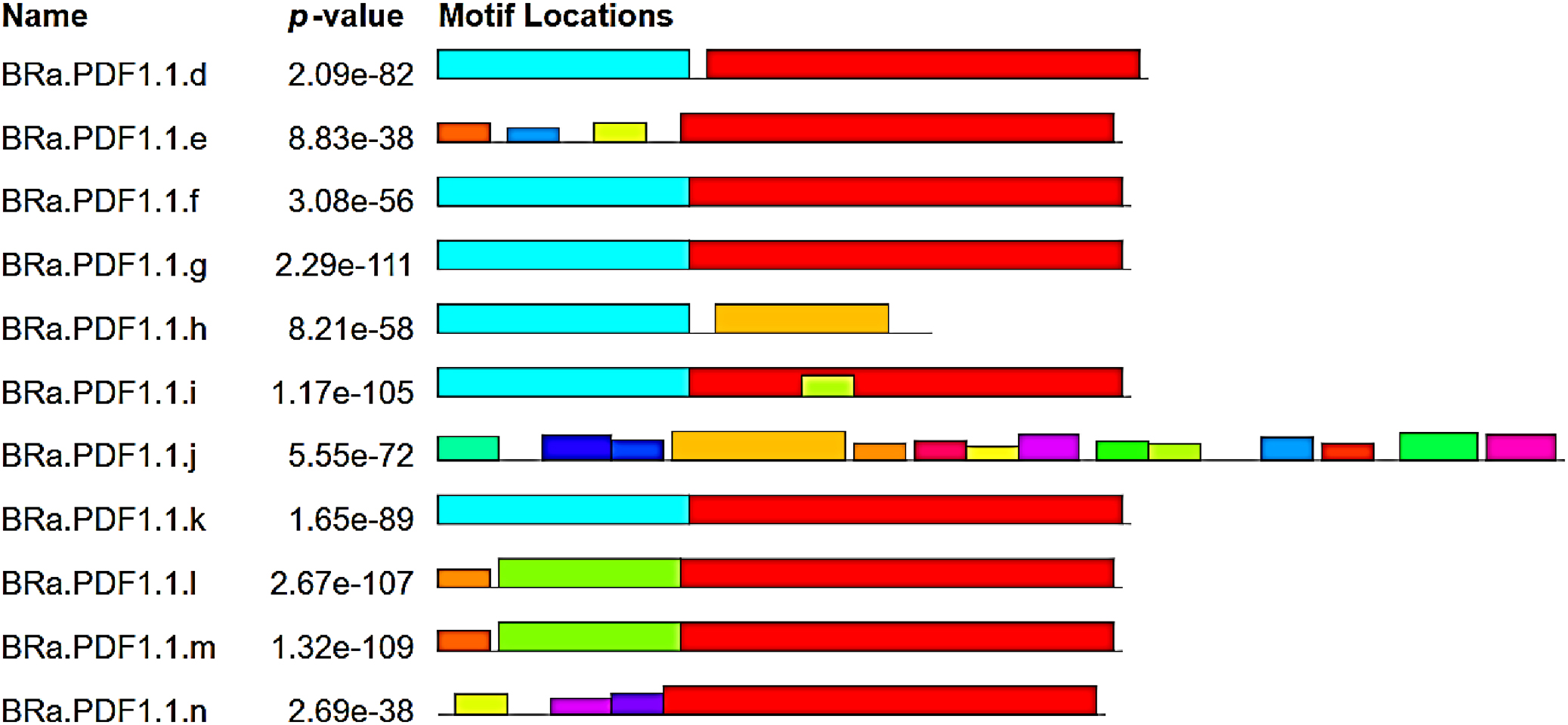
Fig. 7.

Fig. 8.
3.6. Gene ontology (GO) and KEGG analysis
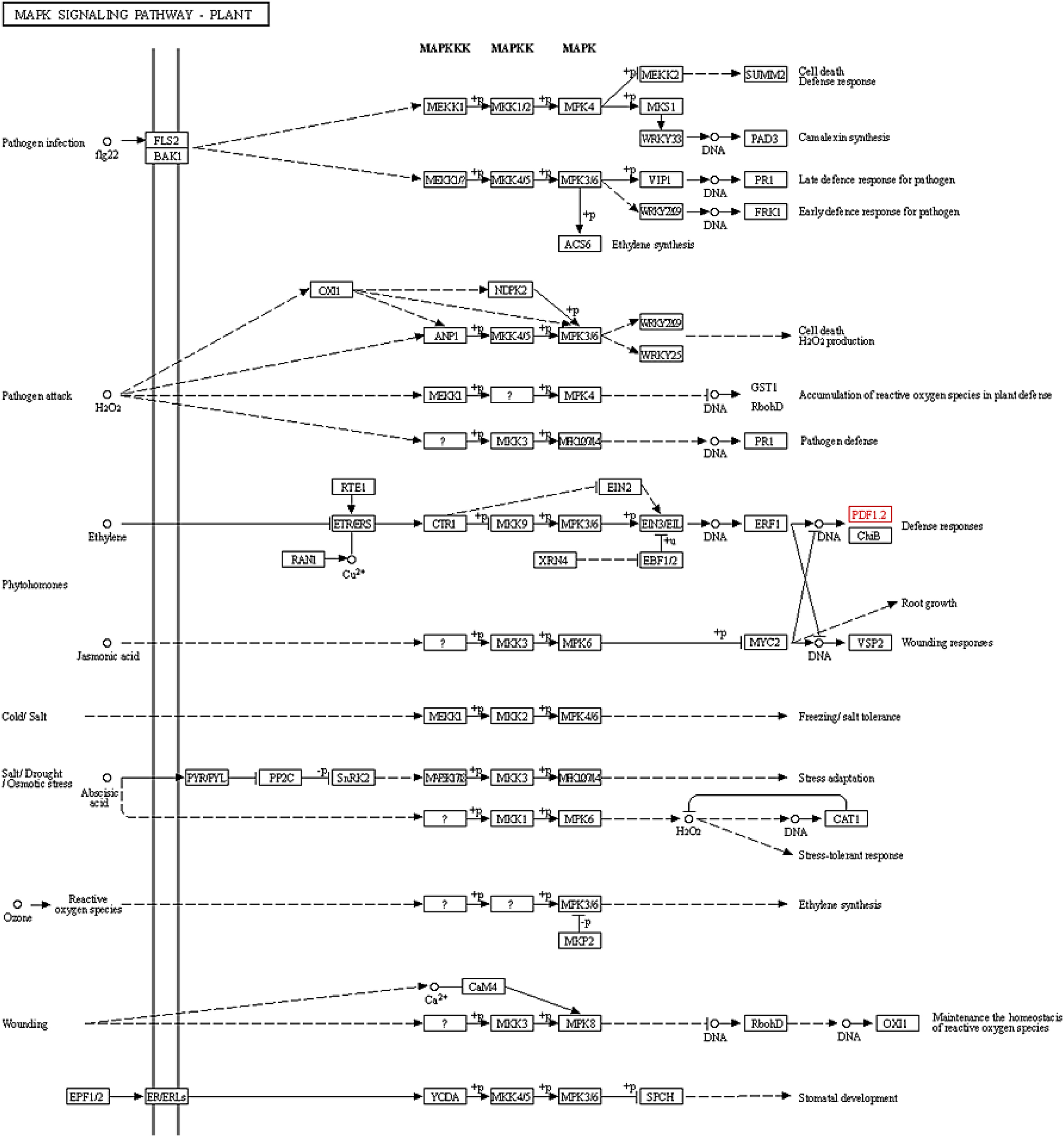
To perform the gene ontology BRa.PDF1.1 proteins, PANZZER2 web server was employed. Nine of 14 proteins were found to be involved in various biological processes. Five proteins were predicted to be involved in different molecular functions and nine proteins were involved in the other cellular processes. The major biological processes found were the degeneration of cells of another organism, defense response to fungus, jasmonic acid, and ethylene-dependent systemic resistance and response to insects (GO:0031640, GO:0050832, GO:0009861, and GO:0009625). Similarly, the molecular function predicted was the protein binding (GO: 0005515), while the cellular components found were extracellular region, secretory vesicle, and membrane (GO: 0005576, GO: 0099503, and GO: 0016020). The KEGG analysis was also performed to explore more functions of Bra.PDF1.1 proteins by BlastKOALA. These results demonstrated that most of the BRa.PDF1.1 proteins were involved in the mitogen-activated protein kinase (MAPK) signaling pathway. The MAPK pathway is involved in the transduction of extracellular signals to the nucleus or cytoplasm for a specific cellular response. In plants, the MAPK signaling pathway is triggered by various biotic or abiotic stress stimuli such as pathogen infection, drought, salinity, high temperature, etc. The pathways in which these proteins were predicted to be involved by KEGG are displayed in Fig. 9.
Fig. 9.
3.7. Prediction of functional sites (N-glycosylation sites and phosphorylation sites)
The protein phosphorylation sites were predicted by using the NetPhos 3.1 server. These results suggested that the proteins had phosphorylation sites ranging from 2 sites to 15 sites with specific and nonspecific kinases. These phosphorylation sites were present only in three specific amino acids, i.e., serine, threonine, and tyrosine. Moreover, no N-Glycosylation sites were predicted in the protein sequences when subjected to the NetNGlyc 1.0 server.
3.8. Secondary structure prediction of Bra.PDF1.1s
The BRa.PDF1.1 protein’s secondary structure was predicted by using the SOPMA online server. Results showed that the alpha helix is the major component of BRa.PDF1.1 proteins. Other respective percentages of all the elements of the secondary structure including the beta sheets, extended strands, random coils, and beta turns are compiled in Table 3.
Table 3.
| Proteins | Alpha helix | Extended strands | Random coils | Beta turns |
|---|---|---|---|---|
| BRa.PDF1.1a | 43.75% | 10.00% | 46.25% | – |
| BRa.PDF1.1b | 40.00% | 15.00% | 43.75% | 1.25% |
| BRa.PDF1.1c | 41.25% | 20.00% | 38.75% | – |
| BRa.PDF1.1d | 41.77% | 11.39% | 43.04% | 3.80% |
| BRa.PDF1.1e | 40.51% | 17.72% | 37.97% | 3.80% |
| BRa.PDF1.1f | 37.50% | 17.50% | 43.75% | 1.25% |
| BRa.PDF1.1 g | 37.50% | 17.50% | 42.50% | 2.50% |
| BRa.PDF1.1 h | 35.37% | 8.54% | 52.44% | 3.66% |
| BRa.PDF1.1i | 41.56% | 11.69% | 41.56% | 5.19% |
| BRa.PDF1.1j | 42.40% | 7.50% | 50.00% | – |
| BRa.PDF1.1k | 37.97% | 16.46% | 41.77% | 3.80% |
| BRa.PDF1.1l | 82.46% | – | 17.54% | – |
| BRa.PDF1.1 m | 12.00% | 32.0% | 45.33% | 10.67% |
| BRa.PDF1.1n | 23.08% | 12.31% | 61.54% | 3.08% |
3.9. Three-dimensional structure and protein pockets prediction
The 3D structures were predicted by using the Phyre (Paul et al. 2019) web server. All the selected protein models represented the maximum confidence (61.1% to 100.0%) and residue coverage (18% to 64%). All of the predicted models are displayed in Table 4. Furthermore, the model quality was evaluated by the Ramachandran plot. This analysis revealed that 59.1% to 100.0% of amino-acid residues in all the proteins under consideration were allocated in the favorable region which suggested the good quality of their predicted structures. The very next was to predict the active sites/pockets of BRa.PDF1.1 proteins. These pockets were predicted by using the CASTp tool. The pockets or active sites that were predicted during this particular analysis are shown in red color in Table 4. The amino-acid residues that were contained in these pockets were Alanine (Ala), Cytosine (Cys), Lysine (Lys), Glutamine (Gln), Isoleucine (Ile), Leucine (Leu), Arginine (Arg), Serine (Ser), Aspartic acid (Asp), Glycine (Gly), Valine (Val), Threonine (Thr), Glutamic acid (Gln), and Tryptophan (Trp). All of these pockets were of different area and volume sizes based on the number of residues they possessed.
Table 4.
4. Discussion
Turnip is also known as B. rapa which is cultivated worldwide. It is also a medicinal plant and has lots of health benefits. Turnips are a rich source of carbohydrates, volatiles, and phenolic compounds, and have many pharmacological benefits such as antioxidant, antidiabetic, and anti-inflammation vegetables (Paul et al. 2019). The growth of B. rapa plant is significantly impacted by various fungal diseases, such as powdery mildew, black spots, and white rust, which collectively contribute to reduced crop yield worldwide. However, PDF1.1 proteins exhibit antifungal properties that inhibit the growth of these harmful fungal pathogens, helping to protect the plant and improve its resilience against these diseases (Cao et al. 2021; Mourou et al. 2023). PDFs are also found in other plants, such as Medicago sativa (alfalfa), where they play a protective role by inhibiting crown rot disease (Sathoff et al. 2019). Our results about the B. rapa PDFs are that these proteins have antifungal properties and are involved in the mitogen-activated protein kinase signaling pathway.
Antimicrobial proteins or peptides including defensins degrade the plasma membrane and pathogens’ cellular activities are impaired (He et al. 2015). PDFs were first studied in wheat and barley seeds and these proteins are localized in the peripheral areas, xylem, or stomata cells (Lacerda et al. 2014). The results of this study showed that these proteins consists of the range of 57 to 130 amino acids (mostly eight cysteine residues) with molecular weights 6162.26 to 13867.87 kDa. These are the stable proteins and its isoelectric point (pI) varies from 5.05 to 9.57. Most of the BRa.PDF1.1 genes from the 14 are located on chromosome no. 7, while some are present on chromosomes no 2, 5, 6, and 8, in B. rapa. Fourteen BRa.PDF1.1 proteins have the same gamma-thionin domain and 20 conserved motifs were identified. We also found that these proteins are localized in extracellular space, vacuoles, nucleus, cytosol, etc. (Zhao et al. 2022). AhDef proteins (peanut defensin) are made up of 66 to 86 amino acids with molecular weight ranging from 7.41 to 9.96 kDa and its isoelectric point (pI) varies from 6.03 to 9.77. Most of the AhDef from 12 genes are located on chromosome no 8 while some are present on chromosomes no 1, 3, 11, 16, and 18. Likewise in BRa.PDF1.1 proteins the AhDef also have a gamma-thionin domain but they have eleven conserved motifs. They also reported in their study these genes are expressed in the plasma membrane or protoplast and these proteins show resistance to Ralstonia Solanacearum bacteria (Vriens et al. 2014). PDFs are small peptides made up of 45 to 54 amino-acid residues and induce their response against biotic or abiotic stress as well as salicylic acid, ethylene, or methyl jasmonate.
Our results regarding the phylogenetic analysis suggested that B. rapa PDF1.1s exhibit maximum homology with Arabidopsis thaliana but also show similarity with peach PDF1.1s in the maximum likelihood tree. The phylogenetic analysis shows that the Brassica napus PDFs (BnaPDFs) show homology with Arabidopsis thaliana PDFs. They also found that they have alpha helix as a main component with 30% to 60%. Gene ontology (GO) of BnaPDFs and BRa.PDF1.1s was the same. The results of BnaPDFs are similar to BRa.PDF1.1s but only BnaA5.PDF1.4 has three exons and two introns. They also report that the BnaPDFs have only one transmembrane except for one protein (BnaC1.PDF1.4). The MEGA X software has also been utilized in other studies, including one focused on generating a neighbor-joining tree to illustrate the evolutionary relationships among MYB proteins from various plants, such as Oryza sativa, Zea mays, B. rapa, Brassica napus, and Arabidopsis thaliana (Luo et al. 2023). The analysis conducted using the Gene Structure Display Server revealed the number and positions of introns and exons within the gene sequences. Through this tool, we discovered that all B. rapa PDF genes contain two exons and a single intron, while B. rapa FPsc.014 specifically exhibits longer exon regions. Additionally, this tool successfully identified the intronic and exonic sequences within chalcone synthase genes, providing further insights into their structural composition (Hussain 2023). Furthermore, gene ontology analysis of Hylocereus polyrhizus using the PANNZER2 web server indicated that its main biological function is the transactivation of downstream genes (Xiao et al. 2025).
The secondary structure analysis of BRa.PDF1.1 proteins revealed that the alpha helix was the dominant structural component, ranging from 12.00% to 82.46%. Additionally, these proteins contained smaller proportions of random coils, beta turns, and extended strands. Similarly, SOMPA was used to predict the secondary structure of the meristem defective protein in the model plant Arabidopsis thaliana, which showed 47.20% alpha helix content (Muccee 2024). The 3D structure predicted via the Phyre (Paul et al. 2019) web server of PDFs has one alpha-helix and three anti-parallel beta-sheets (Lacerda et al. 2014) which is similar to the BRa.PDF1.1s. In contrast, PDFs of Medicago spp, Heuchera sanguinea, and Nicotiana alata are also involved in the MAP kinase signaling pathway (Aerts et al. 2011; Vriens et al. 2014; Dracatos et al. 2016) just like the PDFs of B. rapa. The role of the MAPK signaling pathway has been investigated in sugarcane to understand its response to pathogenic attacks, specifically by the fungus Colletotrichum falcatum, which is the primary causative agent of red rot disease (Gujjar et al. 2024). This disease poses a significant threat to sugarcane crops, impacting yield and quality. By examining the MAPK signaling pathway, researchers aim to uncover the molecular mechanisms involved in sugarcane’s defense against this pathogen (Gujjar et al. 2024). Sclerotinia stem rot caused the Sclerotinia sclerotiorum phytopathogen which affects 400 different hosts/species including the Brassica napus (Bolton et al. 2006). PDFs play a crucial role in a plant’s innate immune system, serving multiple protective functions. Beyond directly destroying fungal pathogens, they inhibit plant cell ion channels, block alpha-amylase and trypsin enzymes, which disrupts insect digestion, and exhibit strong antibacterial properties as well. These combined actions enhance the plant’s defense against a wide range of biotic threats.
5. Conclusions
Brassica rapa (turnip) is a beneficial vegetable grown worldwide and has many good health effects it somehow protects humans from cancer due to the presence of isothiocyanates it also protects humans from diabetes and has many other benefits. Their growth is inhibited by the fungal disease but PDFs 1.1 and in turnips BRa.PDF1.1 are the proteins that overcome this problem due to their anti-fungal activity these proteins are involved in the mitogen-activated-protein-kinase signaling pathway which is triggered by pathogen attack or biotic or abiotic stress, then these proteins kill the pathogenic cells and produce the defense response in B. rapa.
References
Aerts AnM., Bammens L., Govaert G., Carmona-Gutierrez D., Madeo F., Cammue BP.A., Thevissen K. 2011. The antifungal plant defensin HsAFP1 from Heuchera sanguinea induces apoptosis in Candida albicans. Frontiers in Microbiology, 2, 47.
Angamuthu K., Piramanayagam S. 2017. Evaluation of in silico protein secondary structure prediction methods by employing statistical techniques. Biomedical and Biotechnology Research Journal (BBRJ), 1, 29.
Bolton M.D., Thomma B.P., Nelson B.D. 2006. Sclerotinia sclerotiorum (Lib.) de bary: biology and molecular traits of a cosmopolitan pathogen. Molecular Plant Pathology, 7, 1–16.
Cao Q., Wang G., Peng Y. 2021. A critical review on phytochemical profile and biological effects of turnip (Brassica rapa L.). Frontiers in Nutrition, 8, 721733.
Cao Y., Geddes T.A., Yang J.Y.H., Yang P. 2020. Ensemble deep learning in bioinformatics. Nature Machine Intelligence, 2, 500–508.
Chao J., Li Z., Sun Y., Aluko O.O., Wu X., Wang Q., Liu G. 2021. MG2C: a user-friendly online tool for drawing genetic maps. Molecular Horticulture, 1, 1–4.
Chen C., Xia R. 2022. Integrative bioinformatics: history and future. Springer. pp. 343–375.
Dejanovic GM., Asllanaj E., Gamba M., Raguindin P.F., Itodo O.A., Minder B., et al. 2021. Phytochemical characterization of turnip greens (Brassica rapa ssp. rapa): a systematic review. PLoS ONE, 16, e0247032.
Dracatos P.M., Payne J., Di Pietro A., Anderson M.A., Plummer K.M. 2016. Plant defensins NaD1 and NaD2 induce different stress response pathways in fungi. International Journal of Molecular Sciences, 17, 1473.
Garg V.K., Avashthi H., Tiwari A., Jain P.A., Ramkete P.W.R., Kayastha A.M., Singh V.K. 2016. MFPPI–multi FASTA ProtParam interface. Bioinformation, 12, 74.
Gujjar R.S., Kumar R., Goswami S.K., Srivastava S., Kumar S. 2024. MAPK signaling pathway orchestrates and fine-tunes the pathogenicity of Colletotrichum falcatum. Journal of Proteomics, 292, 105056.
He Y., Cao X., Li K., Hu Y., Chen Y-Ru, Blissard G., et al. 2015. A genome-wide analysis of antimicrobial effector genes and their transcription patterns in Manduca sexta. Insect Biochemistry and Molecular Biology, 62, 23–37.
Hu Bo, Jin J., Guo An-Y, Zhang He, Luo J., Gao Ge 2015. GSDS 2.0: an upgraded gene feature visualization server. Bioinformatics, 31, 1296–1297.
Hussain S. 2023. Bioinformatics-based characterization of the Chalcone synthase (CHS) family genes in flowering plants. SABRAO Journal of Breeding and Genetics, 55, 1950–1962.
Jiang Y., Wang D., Wang W., Xu D. 2021. Computational methods for protein localization prediction. Computational and Structural Biotechnology Journal, 19, 5834–5844.
Joshi H.J., Gupta R. 2015. Eukaryotic glycosylation: online methods for site prediction on protein sequences. Glycoinformatics, 127–137.
Kanehisa M., Sato Y. 2020. KEGG Mapper for inferring cellular functions from protein sequences. Protein Science, 29, 28–35.
Kovaleva V., Bukhteeva I., Kit O.Y., Nesmelova I.V. 2020. Plant defensins from a structural perspective. International Journal of Molecular Sciences, 21, 5307.
Lacerda A.F., Vasconcelos É.A., Pelegrini P.B., Grossi de Sa M.F. 2014. Antifungal defensins and their role in plant defense. Frontiers in Microbiology, 5, 116.
Laskowski R.A., Furnham N., Thornton J.M. 2013. Biomolecular forms and functions: a celebration of 50 years of the ramachandran map. World Scientific. pp. 62–75.
Letunic I., Bork P. 2021. Interactive Tree of Life (iTOL) v5: an online tool for phylogenetic tree display and annotation. Nucleic Acids Research, 49, W293–W296.
Li F., Deng Y., Yamamoto E., Liu Z. 2023. 20 Plant Omics Databases: an Online Resource Guide.
Li F., Li C., Marquez-Lago T.T., Leier A., Akutsu T., Purcell A.W., et al. 2018. Quokka: a comprehensive tool for rapid and accurate prediction of kinase family-specific phosphorylation sites in the human proteome. Bioinformatics, 34, 4223–4231.
Luo D., Mei D., Wei W., Liu J. 2023. Identification and phylogenetic analysis of the R2R3-MYB subfamily in Brassica napus. Plants, 12, 886.
Mourou M., Raimondo M.L., Lops F., Carlucci A. 2023. Brassicaceae fungal diseases: molecular detection and host–plant interaction. Plants, 12, 1033.
Muccee F. 2024. In-silico characterization of meristem defective (MDF) protein associated with regulation of root meristems in Arabidopsis thaliana. Pakistan Journal of Science, 76, 76–83.
Nystrom S.L., McKay D.J. 2021. Memes: a motif analysis environment in R using tools from the MEME Suite. PLoS Computational Biology, 17, e1008991.
Paul S., Geng C.A., Yang T.H., Yang Y.P., Chen J.J. 2019. Phytochemical and health-beneficial progress of turnip (Brassica rapa). Journal of Food Science, 84, 19–30.
Sathoff A.E., Velivelli S., Shah D.M., Samac D.A. 2019. Plant defensin peptides have antifungal and antibacterial activity against human and plant pathogens. Phytopathology®, 109, 402–408.
Stecher G., Tamura K., Kumar S. 2020. Molecular evolutionary genetics analysis (MEGA) for macOS. Molecular Biology and Evolution, 37, 1237–1239.
Sun X., Wang W., Paerhati M., Song Z., Li J., Zhu J. 2022. Phytochemical composition and nutritional characterization of qamgur (Brassica rapa L.) in different forms. Food Science and Technology, 42.
Tian W., Chen C., Lei X., Zhao J., Liang J. 2018. CASTp 3.0: computed atlas of surface topography of proteins. Nucleic Acids Research, 46, W363–W367.
Törönen P., Holm L. 2022. PANNZER—a practical tool for protein function prediction. Protein Science, 31, 118–128.
Vriens K., Cammue B.P., Thevissen K. 2014. Antifungal plant defensins: mechanisms of action and production. Molecules (Basel, Switzerland), 19, 12280–12303.
Xiao L., Cai X., Yu R., Nie X., Wang Ai-H, Yang K., Wen X. 2025. Genome-wide identification of Pitaya (Hylocereus polyrhizus) TCPs and their involvement in flower development and abiotic stress response. Tropical Plant Biology, 18, 9.
Yang M., Derbyshire MK., Yamashita RA., Marchler‐Bauer A. 2020. NCBI's conserved domain database and tools for protein domain analysis. Current Protocols in Bioinformatics, 69, e90.
Yao J., Luo J.-S., Xiao Y., Zhang Z. 2019. The plant defensin gene AtPDF2. 1 mediates ammonium metabolism by regulating glutamine synthetase activity in Arabidopsis thaliana. BMC Plant Biology, 19, 1–13.
Yao J., Luo J.-S., Xiao Y., Zhang Z. 2019. The plant defensin gene AtPDF2. 1 mediates ammonium metabolism by regulating glutamine synthetase activity in Arabidopsis thaliana. BMC Plant Biology, 19, 1–13.
Zhao K., Ren R., Ma X., Zhao K., Qu C., Cao Di, et al. 2022. Genome-wide investigation of defensin genes in peanut (Arachis hypogaea L.) reveals AhDef2. 2 conferring resistance to bacterial wilt. The Crop Journal, 10, 809–819.
Supplementary material
Supplementary Material 1 (DOCX / 20 KB).
- Download
- 19.31 KB
Information & Authors
Information
Published In

FACETS
Volume 10 • 2025
Pages: 1 - 16
Editor: Vance L Trudeau
History
Received: 1 March 2024
Accepted: 8 December 2024
Version of record online: 11 March 2025
Copyright
© 2025 The Author(s). This work is licensed under a Creative Commons Attribution 4.0 International License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Data Availability Statement
Data generated or analyzed during this study are provided in full within the published article.
Key Words
Sections
Subjects
Authors
Author Contributions
Conceptualization: QN, MFS
Data curation: MFA, IA
Formal analysis: MFS, MFA, SA, IM, YS
Funding acquisition: IA
Investigation: QN
Methodology: QN
Project administration: QN, MFS, SA, YS, IA
Resources: MFA, IM, YS, IA
Software: QN, YS
Supervision: MFS, SA
Validation: MFS, SA, IM
Visualization: MFA, IM, YS, IA
Writing – original draft: QN
Writing – review & editing: MFS
Competing Interests
The authors declare there are no competing interests.
Funding Information
The authors are thankful to the Deanship of Research and Graduate Studies, King Khalid University, Abha, Saudi Arabia, for financially supporting this work through the Large Research Group Project under Grant no. R.G.P2/513/45.: R.G.P2/513/45
The authors are thankful to the Deanship of Research and Graduate Studies, King Khalid University, Abha, Saudi Arabia, for financially supporting this work through the Large Research Group Project under Grant No. R.G.P2/513/45.
Metrics & Citations
Metrics
Other Metrics
Citations
Cite As
Qudsia Naz, Muhammad Farhan Sarwar, Mudassar Fareed Awan, Sajed Ali, Irfan Mughal, Yousuf Shafiq, and Irfan Ahmad. 2025. Multifacets in silico study of Plant Defensin 1.1 protein (PDF1.1) in Brassica rapa (turnip). FACETS.
10: 1-16.
https://doi.org/10.1139/facets-2024-0038
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
There are no citations for this item