The decline of walleye populations: an ecological tipping point?
Abstract
Walleye/ogaa (Sander vitreus (Mitchill)) (hereafter, walleye; ogaa = Ojibwe translation) populations have historically supported important multi-use, harvest-oriented fisheries. Despite intensive management, walleye populations have declined in the midwestern United States raising concerns about the sustainability of the species. Numerous factors have been implicated in walleye population declines, including climate change, habitat loss, invasive species, species-interactions, production overharvest (i.e., harvest consistently exceeding annual production), and changing angler behaviors. These factors have negatively influenced natural recruitment and contributed to depensatory recruitment dynamics. I provide a review and perspective suggesting that the current trajectory of walleye populations is at or nearing an ecological tipping point. Although fish populations are often considered compensatory (i.e., negatively density-dependent), current walleye populations appear prone to depensation (i.e., positive density dependence). My review and perspective suggest that a compensatory management perspective for walleye is misaligned. A change in management towards a depensatory resource focus using ecosystem-based fisheries management and the recognition of walleye fisheries as social–ecological systems is needed for conservation. If compensatory management ensues, walleye persistence will likely be further threatened because many drivers of change are outside of managerial control, and those commonly used within managerial control have seemingly been ineffective for sustaining or rehabilitating naturally reproducing walleye populations.
Introduction
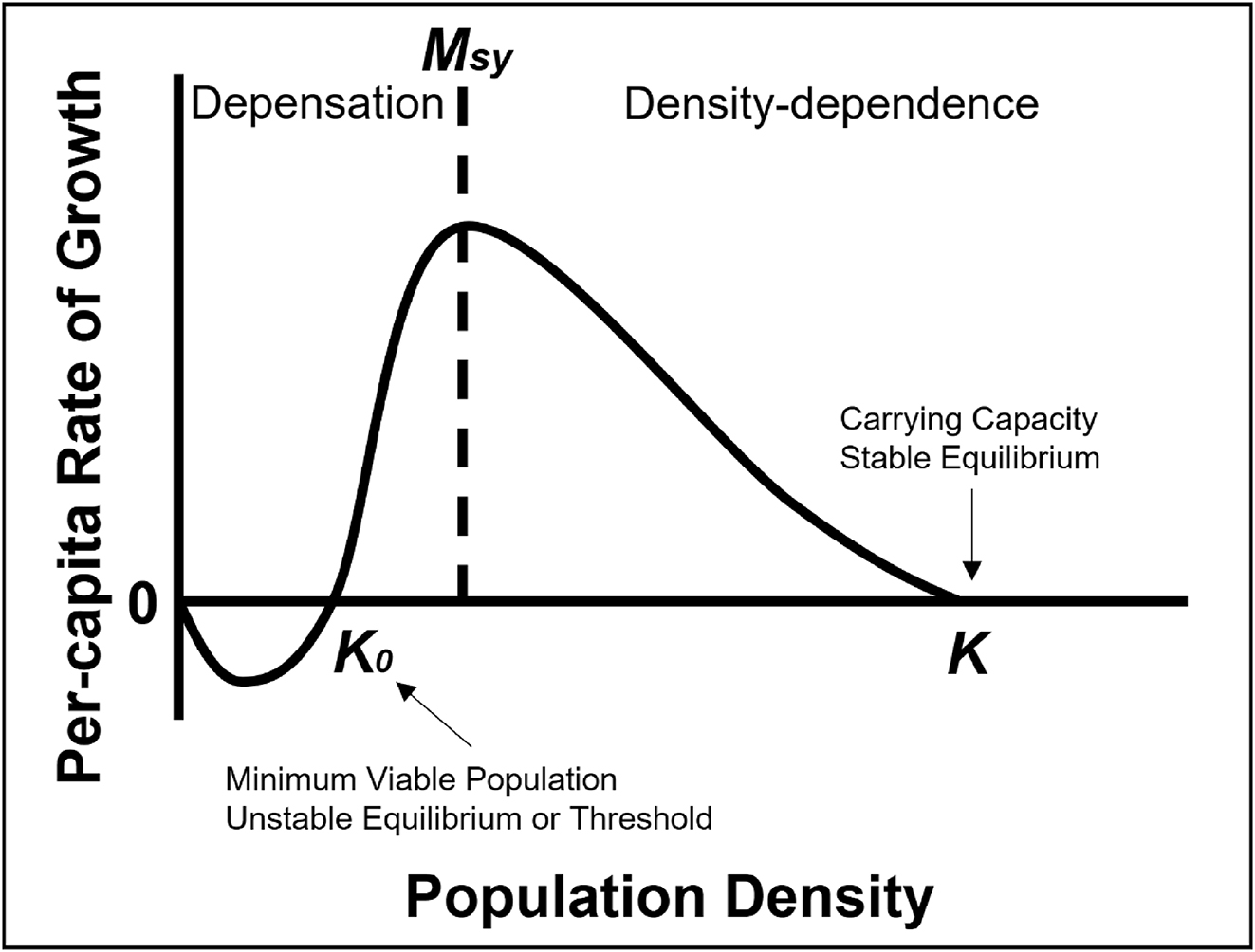
Exploited fish populations have long been managed as compensatory (i.e., negatively density-dependent) resources through various input (i.e., limitations on fishing effort) and output controls (i.e., limits on the number of fish coming out of a fishery) (Bellido et al. 2020). Common input controls have included harvest quotas or seasonal fishery closures. Input controls have been used less extensively for recreational fisheries, whereas they are common in high-efficiency commercial and subsistence fisheries. Output controls are common to recreational fisheries where length, bag, and possession limits create restrictions on the length, number, and possession of fish harvested. Both regulation types are aimed to limit fisher effort to various degrees and to manage harvest to prevent overexploitation. Output controls may be less effective for limiting fisher effort and in preventing overexploitation in open access fisheries. Exploited fisheries are assumed to be resilient to harvest through density-dependent survivorship of recruits as adult stock sizes decline (Ricker 1954). However, density-dependent recruit survival may be compromised when fisher effort and efficiency remain high, despite low adult stock sizes and depensatory thresholds leading to population collapse are crossed (Liermann and Hilborn 1997, 2001; Walters and Kitchell 2001; Post 2013; Sass et al. 2021; Dassow et al. 2023) (Fig. 1). It is important to note that fishery-induced stressors are not the only variables influencing fish population sustainability that can result in depensatory (i.e., positively density-dependent) recruitment dynamics (Walters and Kitchell 2001; Post et al. 2002; Sass et al. 2021; Dassow et al. 2023). Because fisheries have collapsed or have been slow to recover following drastic management intervention (e.g., fishery closure), assessment of long-standing fisheries management paradigms and assumptions may need to be revisited and reimagined based on current scientific knowledge. Then, accordingly adapted to responsibly manage and conserve exploited fisheries now, and in the future, to avoid repeating the failures of the past. Failure to adapt to changing environmental and social conditions influencing fisheries risks treating exploited fish populations as infinitely compensatory resources or ones where human ingenuity and engineering (e.g., aquaculture, stocking, artificial habitat modification of systems) is even more heavily relied upon to conserve fish populations and allow for harvest opportunity (Lorenzen 2014). The latter approach assumes that “engineering solutions” will always overcome abiotic and biotic factors negatively influencing harvested fish populations.
Fig. 1.
Walleye/ogaa (Sander vitreus (Mitchill)) (hereafter, walleye) in North America provide a model example of a fish that has been intensively managed due to the harvest-oriented nature of recreational and subsistence fisheries (Barton 2011; Gaeta et al. 2013; Hansen et al. 2015). Walleye populations support culturally, economically, and recreationally important fisheries throughout their native and introduced range. In the midwestern United States and portions of Canada, walleye populations are exploited by Native American or First Nations subsistence fishers and recreational anglers, and their fisheries are often embedded in complex social–ecological systems (Fig. 2). In these joint subsistence and recreational walleye fisheries, high efficiency subsistence fisheries are generally managed using input controls (e.g., quotas) and lower efficiency recreational fisheries are primarily managed using output controls (e.g., length, bag, and possession limits) (U.S. Department of the Interior 1991; Hansen et al. 2005; Mrnak et al. 2018). Despite intensive management, many walleye populations have declined over time due to persistent natural recruitment failures and sometimes overharvest (Post et al. 2002; Embke et al. 2019; Cahill et al. 2022) (Fig. 2).
Fig. 2.
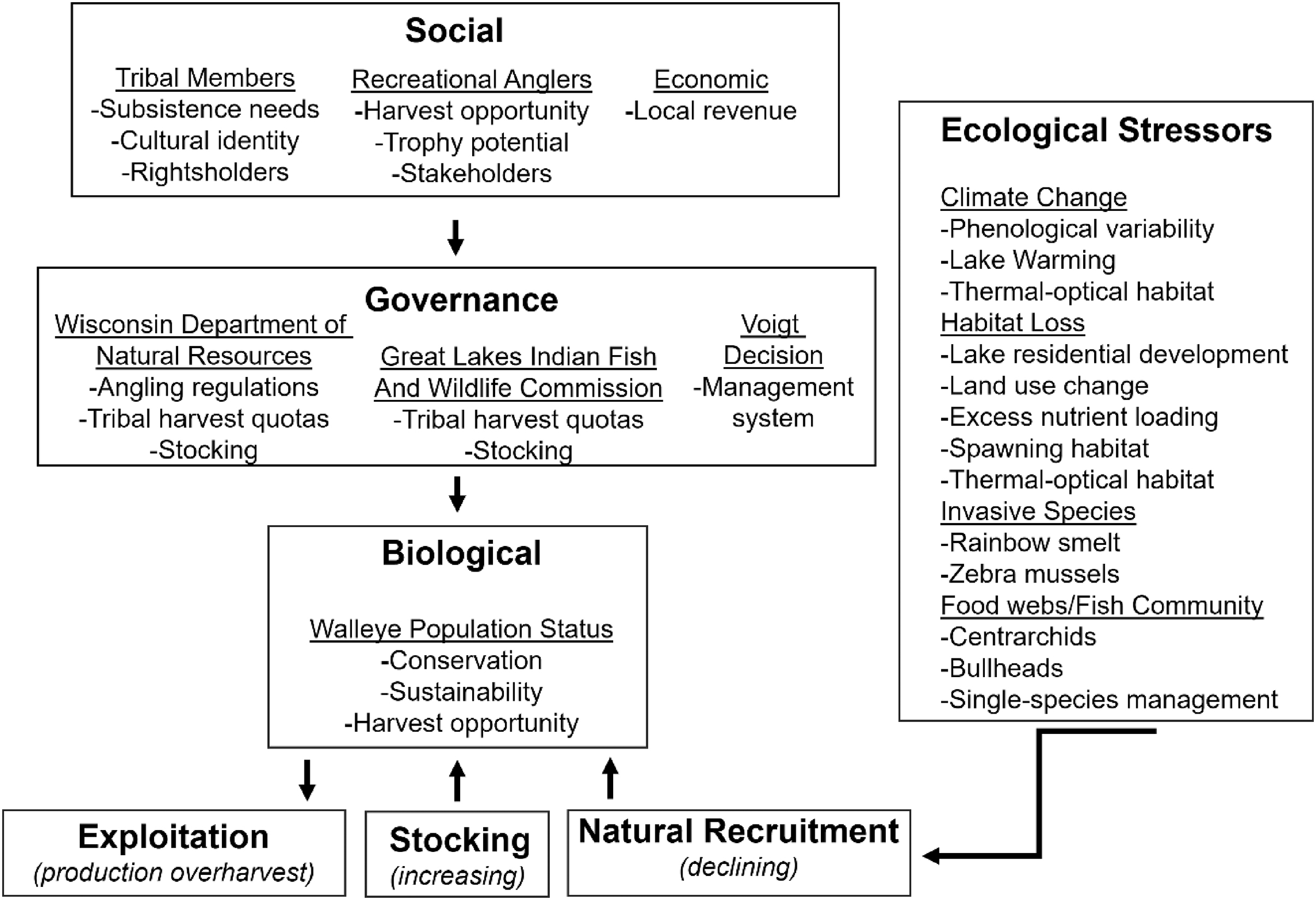
Sustainability of exploited fish populations is dependent on consistent natural recruitment, which allows for stock replenishment and harvest opportunities of surplus production (Ricker 1975). Walleye natural recruitment has declined or failed in many systems within the species native range of the midwestern United States over time (Embke et al. 2019). Walleye natural recruitment declines and failures have resulted in reductions in adult abundance, biomass, production, and production:biomass ratios (i.e., population replacement rate) (Hansen et al. 2015; Pederson et al. 2018; Rypel et al. 2018; Embke et al. 2019; Mrnak et al. 2024). Walleye natural recruitment declines have prompted extensive stocking of the species at various life stages (e.g., fry, small fingerling, large fingerling) as one measure to combat reductions in population abundance and to reestablish natural recruitment (Lawson et al. 2022; Sass et al. 2022a; Elwer et al. 2023) (Fig. 2). Several abiotic, anthropogenic, and biotic factors (and their interactions) have been implicated in walleye natural recruitment declines. Slow and abrupt environmental change of abiotic factors influencing natural recruitment have included directional climate change and associated lake warming (Hansen et al. 2017, 2018; Feiner et al. 2022a), climate change-induced phenological variability in lake ice dynamics (Feiner et al. 2022a; Barta et al. 2024), changes in available thermal–optical habitat (Lester et al. 2004; Mahlum et al. 2023), and land use, watershed, and within-system habitat change (Sass et al. 2017; Dassow et al. 2023) (Fig. 2). Production overharvest (i.e., harvest consistently exceeding annual production) of walleye populations has also been observed in northern Wisconsin lakes (Embke et al. 2019; Mrnak et al. 2024) (Fig. 2). Production overharvest has occurred because exploitation by the recreational and subsistence fisheries has been stable or increasing, while natural recruitment and associated production replacement rates (production:biomass ratios) have declined over time (Rypel et al. 2018; Embke et al. 2019; Mrnak et al. 2024). Walleye natural recruitment has also been negatively associated with fish community composition change over time, particularly increases in centrarchids such as largemouth bass (Micropterus salmoides (Lacepede)) (Hansen et al. 2015; Hansen et al. 2017, 2018; Dassow et al. 2023), black crappie (Pomoxis nigromaculatus (Lesueur)) (Broda et al. 2022), and bluegill (Lepomis macrochirus Rafinesque) (Embke et al. 2022) (Fig. 2). High voluntary angler release rates of largemouth bass and panfish, along with conservative harvest regulations in these fisheries, have also contributed to abundance increases of centrarchids over time (Gaeta et al. 2013; Hansen et al. 2015; Rypel 2015; Sass and Shaw 2020; Sass et al. 2023). Overabundant bullhead Ameiurus spp. populations have also been shown to negatively influence walleye natural recruitment (Sikora et al. 2021) (Fig. 2). Last, empirical evidence has suggested that walleye are prone to depensatory recruitment dynamics (Sass et al. 2021; Dassow et al. 2023). Mechanisms increasing the probability of depensatory recruitment in walleye have been linked to interactions among largemouth bass relative abundance, climate, and land use change (Dassow et al. 2023).
My review and perspective aim to discuss traditional assumptions and strategies used to manage exploited fisheries under the auspice that these approaches may lack applicability and relevance now, and in the future, to conserve fish populations and provide harvest opportunity. My perspective is based on first principles of natural resource management: (1) sustaining natural resources in perpetuity, and (2) if (1) is met, allowed for harvest opportunity in fisheries. My review and perspective focus on North American walleye fisheries, although the subject matter should be broadly applicable to other exploited fish populations. The specific objectives of my review and perspective include: (1) discussion of current walleye management approaches and justifications; (2) evaluation of fisheries management assumptions and paradigms for managing exploited walleye fisheries and outcomes; and (3) alternative courses of action to conserve walleye populations when current fisheries assumptions and paradigms may fail to contribute to long-term, sustainable outcomes. My review and perspective consider the biology and ecology of walleye, human dimensions of walleye fisheries, and is largely based on ecosystem-based fisheries management, fisheries as social–ecological systems, and resilience theory.
Walleye management systems, paradigm shifts, and potential courses of action
Input controls
Input controls (i.e., limitations on fishing effort) used to manage walleye generally consist of closed spring fishing seasons in recreational fisheries (e.g., first Sunday in March to first Saturday in May in Wisconsin lakes), quotas in some recreational fisheries (e.g., Mille Lacs, Minnesota), and the seasonality of tribal subsistence fisheries that primarily focus on spawning walleye in spring. Most waterbodies in the midwestern United States and Canada are open access, meaning that recreational angler effort is not regulated outside of closed fishing seasons. Although open access fisheries are one of the greatest privileges afforded to anglers, the current trajectory of walleye populations in some regions may require consideration of increased angler effort limitation, including additional or longer closed seasons, gear restrictions, and (or) harvest quotas to conserve diminished walleye resources. In extreme circumstances, a lottery system could be used to regulate angler effort and exploitation on individual waterbodies. Effort limitation has been proposed in British Columbia rainbow trout (Oncorhyncus mykiss (Walbaum)) fisheries (Cox et al. 2002), is used in a Wisconsin lake sturgeon (Acipenser fulvescens Rafinesque) spearfishery (Bruch 1999), and is relatively common for hunting wildlife on public and private lands (Boxall 1995; Scrogin et al. 2000). Walleye recreational fisheries are typically closed in late winter/early spring to protect mature females. Angler catch and harvest rates of walleye typically peak in late spring/early summer (May, June), with another minor peak in fall (Bailey et al. 2019; Sass et al. 2023). Additional or prolonged closed seasons during high angler catch and harvest rate periods could be used to conserve the species in dire situations. The use of artificial baits by walleye anglers has also been shown to reduce catch and harvest rates and effort per trip compared to live bait users (Bailey et al. 2019). Artificial bait-only regulations could serve to reduce exploitation and walleye angler effort. Although more common in tribal subsistence fisheries for walleye due to high efficiency (Hansen et al. 2005; Mrnak et al. 2018), harvest quotas could be used in recreational fisheries to protect walleye populations. Harvest quotas in recreational walleye fisheries would require angler creel surveys to track annual harvest. Such measures are currently being used on Mille Lacs, Minnesota, and in a walleye production overharvest experiment on Escanaba Lake, Wisconsin (Sass et al. 2022b), and are certainly not outside the realm of possibility for agencies considering tradeoffs between supporting fisheries-dependent or fisheries-independent assessments of fish populations. That said, it has clearly been established that fisheries-dependent data alone are inadequate for assessing fish population status due to hyperstability in angler catch rates for many fishes (Hansen et al. 2005; Ward et al. 2013; van Poorten et al. 2016; Mrnak et al. 2018; Dassow et al. 2020; Feiner et al. 2020a; Mosley et al. 2022). Management agencies must ponder the tradeoffs of using limited resources and funding to best assess walleye population status through fisheries-independent surveys, fisheries-dependent creel surveys, or both. Angler receptiveness of walleye harvest quotas and communication of harvest periods ending may be inconvenient for anglers and management agencies but seem a small price to pay for tradeoffs between harvest opportunity and fisheries collapse. Collectively, input controls such as lottery systems, fishery closures during highly vulnerable periods, gear restrictions, and harvest quotas could be used in circumstances where the probability of fishery collapse (i.e., nearing an ecological tipping point or depensatory threshold) is high.
Output controls
Output controls (i.e., limits on the number of fish coming out of a fishery) have long been used to prevent overexploitation of walleye populations and to meet population-specific demographic objectives and angler desires (e.g., size-structure, trophy potential). Minimum length limits in recreational fisheries have generally corresponded with length at maturity. For many walleye populations, this roughly corresponds with 381 mm minimum length limits or walleye ranging in age from about 3 to 5 due to sexually dimorphic growth (Sass 2001; Henderson et al. 2003; Sass and Kitchell 2005; Rennie et al. 2008). Tribal subsistence fisheries targeting walleye during spring spawning generally harvest mature fish (mean length = 398 mm; Mrnak et al. 2018). Daily bag limits for walleye in recreational angling fisheries in the midwestern United States and Canada generally range from 2 to 6 walleye/angler/day, with more liberal daily bag limits associated with greater population abundance and consistent natural recruitment, and more conservative regulations associated with lower population abundance or allocations of harvest between recreational and subsistence fisheries. Possession limits for walleye are typically double the daily bag limit.
Given the harvest-oriented nature of walleye fisheries (Gaeta et al. 2013; Hansen et al. 2015), known knife-edge vulnerabilities of walleye >381 mm (Tsehaye et al. 2016), and high angler effort and high-efficiency subsistence fisheries (Mrnak et al. 2018), I am continually baffled by minimum length limits that target the harvest of recently maturing fish. Effectiveness of such regulations rely on consistent natural recruitment to replenish adult stocks and to provide harvest opportunity. Using this management strategy, walleye may only have one or two opportunities to spawn prior to being vulnerable to harvest. This strategy seems non-sensical and the antithesis of precautionary given that environmental factors have been consistently shown to be better predictors of age-0 walleye recruitment than adult stock size (Hansen et al. 1998; Beard et al. 2003; Shaw et al. 2018). Such strategies may challenge walleye population sustainability when natural recruitment is inconsistent, and “mismatches” exceed “matches” in recruitment success (Cushing 1990; Feiner et al. 2022a; Barta et al. 2024). Maternal-related effects (i.e., proportion of longer female walleye in a population, body condition) have also been shown to be better predictors of age-0 recruitment than adult stock size (Shaw et al. 2018; Feiner et al. 2019). No harvest slot length limits (e.g., 508–620 mm) have been used to protect longer female walleye in recreational fisheries, and daily bag restrictions on the number of fish greater than a certain length (e.g., 559 mm) have been used in subsistence fisheries for the same reason. Given current angling regulations, subsistence fisheries targeting recently maturing walleye, and knife-edge vulnerability, no harvest slot length limits may do little to protect longer females within walleye populations. Minimum length limits are enacted to limit exploitation, as are daily bag and possession limits. Nevertheless, daily bag limits may be ineffective for reducing exploitation in open access fisheries subject to high angler effort and low regulatory compliance (Sullivan 2002). This is particularly true for walleye fisheries where catch and harvest rates in angling and subsistence fisheries have been shown to be hyperstable across a range of adult stock sizes and productivities (Hansen et al. 2005; Mrnak et al. 2018, 2024; Mosley et al. 2022). Likewise, possession limit infractions may be difficult to enforce given that “just cause” or “reasonable doubt” is often required, and conservation officer funding and personnel are often limited in waterbody-rich landscapes.
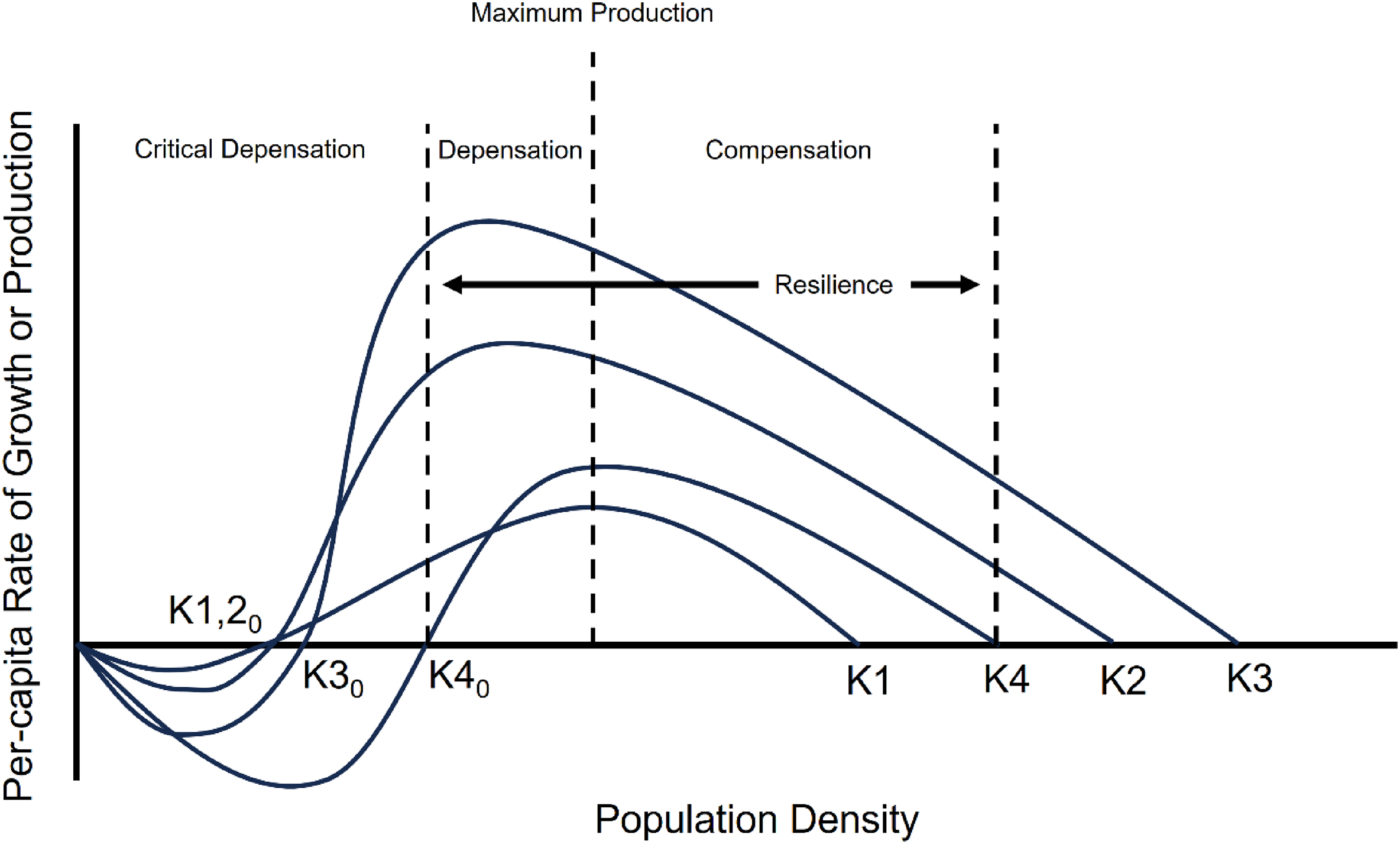
Length, bag, and possession limits may be providing little protection to walleye populations struggling with inconsistent natural recruitment. Current policies have and will likely continue to lead to diminished adult population abundance and production (Hansen et al. 2015; Rypel et al. 2018; Embke et al. 2019; Mrnak et al. 2024), even with intensive walleye stocking and rehabilitation efforts (Lawson et al. 2022; Sass et al. 2022a; Elwer et al. 2023). I propose that input controls should be more stringent in declining walleye populations with inconsistent natural recruitment or perhaps for all walleye populations within a region given current and projected walleye natural recruitment, adult abundance, and production trajectories. At the least, management agencies may consider ≥457 mm minimum length limits and reductions in daily bag (≤3 fish/day) and possession limits to further reduce exploitation and be protective of adult fish. Although adult stock size has been shown to be a weak predictor of age-0 walleye recruitment, zero adults equal zero recruits. Further, reductions in adult walleye stock size >0 may lead to depensatory recruitment (i.e., ecological tipping point, increased probability of population collapse without intervention) and adult abundance thresholds for depensation likely differ among walleye populations (Sass et al. 2021; Dassow et al. 2023) (Fig. 3). Stocking interventions to maintain adult walleye abundance above depensatory thresholds have shown little promise for rehabilitating natural recruitment and sustaining populations (Grausgruber and Weber 2021a, 2021b; Sass et al. 2021; Lawson et al. 2022; Dassow et al. 2023; Elwer et al. 2023). In struggling walleye populations, more conservative harvest regulations afford mature fish multiple opportunities to spawn. Anecdotal management of walleye through observations of a “good” year class every 4–5 years to support a fishery is unreliable (Rypel et al. 2018), and abiotic drivers of recruitment are changing and more variable. Given this, the only recourse is to give mature fish more opportunity for a “match” in recruitment by further reducing exploitation rates (Feiner et al. 2022a; Barta et al. 2024).
Fig. 3.
Stocking
Walleye have a long history of being stocked into waterbodies with natural recruitment, to introduce walleye to create self-sustaining populations (Sass et al. 2022a), and for put–grow–take fishing opportunities. Hatchery products have typically consisted of fry (stocked shortly after eye up), small fingerlings (stocked in mid-summer), and more recently large fingerlings (stocked in fall). Due to reduced production costs and number, fry and small fingerlings were more commonly stocked in early attempts—and still are to create stocked year classes (i.e., overwhelm a system with high numbers of stocked walleye to create a year class despite high mortality). Due to variable success of fry and small fingerling stockings, large fingerling stockings have been more recently favored based on the assumption of greater survival, contribution to stocked year class strength, and a higher probability of re-establishing natural recruitment in formerly naturally reproducing populations (Grausgruber and Weber 2021a, 2021b; Lawson et al. 2022; Sass et al. 2023; Elwer et al. 2023). Large fingerlings have been presumed to have a higher probability of overcoming fry and small fingerling recruitment bottlenecks (i.e., density-dependent mortality) (Boehm 2016; Gostiaux et al. 2022) despite the higher cost of production (Elwer et al. 2023).
Walleye stocking has rarely contributed meaningfully to walleye population establishment and natural recruitment in recent times (Embke et al. in review). In the mid-1900s, walleye stocking was able to create walleye fisheries in some systems, and even natural recruiting populations that persist to this day (e.g., Escanaba Lake, Wisconsin; Sass et al. 2022b). Many of these populations were also ones that showed minor and short-term periods of walleye natural recruitment, were then categorized as lakes managed for walleye, and are now subject to legacy effects by rights holders, anglers, stakeholder groups, and management agencies as “walleye lakes”. Systems such as Escanaba Lake are the anomaly, not the norm. Legacy influences thus dictate continued stocking and walleye management to appease rights holder, angler, and stakeholder group desires when these lakes may not have been “walleye lakes” to begin with (Kitchell et al. 1977; Raabe et al. 2020; Dassow et al. 2022), and the long-held management panacea of stocking by these groups and the funding to drive hatchery infrastructure (Arlinghaus et al. 2022; Radinger et al. 2023). The scientific literature has clearly shown that walleye stocking has often failed to reach lofty goals of establishing naturally reproducing populations or supplementing such populations. Stocking has achieved low abundance, “fishable” populations, and many agencies continue stocking to support these low-density walleye fisheries to meet fisher desires under the guise of population sustainability or re-establishing natural reproduction. Future research should identify lakes with native walleye populations. In north-central North America, these would be walleye native to lakes following the last glaciation about 10 000 years ago. Lake sediment coring and environmental DNA sampling holds promise for determining lakes with native walleye populations. Identification of native walleye lakes may help managers allocate limited resources and set expectations for tribal members, anglers, and stakeholder groups about lakes incapable of supporting walleye populations or natural recruitment, along with other fish-based lake classification systems and decision–support tools (Rypel et al. 2019; Dassow et al. 2022).
Recent empirical evidence suggested that ecosystem-based fisheries management outperformed stocking for influencing fish production (Radinger et al. 2023). Walleye stocking has consistently failed to enhance year class strength in natural reproducing populations (Li et al. 1996) and to re-establish natural reproduction (Rypel et al. 2018) even in populations predicted to have the highest probability of supporting natural recruitment (Hansen et al. 2015a; Elwer et al. 2023). Numerous issues have also resulted from stocking, including genetic introgression and loss of local adaptation (Lorenzen 2014; Bootsma et al. 2021), poor survivorship due to difficulties transitioning to the natural environment (Grausgruber and Weber 2021a) and transport stress (Grausgruber and Weber 2021b), high cost per individual to harvestable age (Elwer et al. 2023), and skewed sex ratios towards females in large fingerling walleye (Sass et al. 2022a). Despite efforts to increase survivorship of the stocked product by raising longer walleye or attempts to engineer a stocked walleye year class by overwhelming a system with tens of millions of walleye fry, stocking outcomes have still shown poor performance leading to low density walleye populations of poor fishing quality or harvest opportunity or predefined goals of put–grow–take fisheries. Given the lack of success of walleye stocking, conservation of existing naturally recruiting walleye populations is paramount to allow for alternative management to preserve remaining populations.
Stocking and rehabilitation efforts have been used in an attempt to increase walleye population resilience, although not explicitly for this purpose or in those terms (Fig. 3). Instead of consistently exploiting walleye populations, especially given improving fishing technologies (Feiner et al. 2020b; Cooke et al. 2021; but see Neely et al. 2023), exploitation should be reduced to decrease the probability of walleye populations crossing an ecological tipping point or depensatory threshold (Figs. 3 and 4). Responsible management to conserve imperiled fisheries resources is to relieve pressure and stressors on those populations. Important levers in addition to conservative harvest regulations and stocking that could be used to increase walleye population resilience include limiting effort, gear restrictions (Bailey et al. 2019), and the promotion of catch-and-release in angling fisheries. In dire conservation situations, seasonal closures could be extended to limit or eliminate directed walleye effort during vulnerable periods because angler and tribal (spring) effort track these seasonal vulnerabilities (Sass et al. 2023). Likewise, catch-and-release could be promoted for walleye, although this may have consequences for growth, recruitment, and angler desires of trophy potential (Haglund et al. 2016; Sass and Shaw 2020). Collectively, compromise and buy-in for walleye conservation by anglers, rights holders, and stakeholder groups, and more aggressive approaches may be needed to avoid crossing depensatory thresholds for walleye and other exploited fisheries (Fig. 4). The alternatives risk depensation, lack of population recovery, and population extirpation (Fig. 4). Some researchers and managers relying on traditional single-species management tenets of compensatory fisheries resources have postulated that reducing harvest may result in wasted surplus production ending in unnecessary natural mortality (Radomski 2022). For walleye, the scientific evidence suggests that we are well beyond the assumption of compensation (Fig. 4). A loss of limited surplus production to anglers and tribal members in some fisheries and reductions in harvest seem a small price to pay for species conservation.
Fig. 4.
Walleye exploitation
Walleye management in joint tribal subsistence (i.e., spearfishing and gill netting) and recreational angling fisheries generally consist of a combination of closed seasons, output controls, and harvest quotas in the case of tribal subsistence fisheries (U.S. Department of the Interior 1991) (Fig. 2). Seasonal closures are aimed to protect walleye during vulnerable periods (e.g., late winter–early spring, Sass et al. 2023), and angler regulations and tribal harvest quotas are used to manage exploitation rates to a level commensurate with long-term population sustainability. Often, “sustainable” exploitation rates are used to set a limit reference point exploitation rate that should not be frequently exceeded (Staggs et al. 1990; Hansen et al. 1991; Sass et al. 2004; Tsehaye et al. 2016). Such walleye management systems rely upon a compulsory creel census of tribal subsistence harvest and point-intercept angler creel surveys and mark–recapture population estimates to determine annual recreational angler exploitation rates to assure non-exceedance of the limit reference point exploitation rate. In the joint walleye fishery in the Ceded Territories of Wisconsin, the limit reference point exploitation rate is 35% of an adult population, with safety factors used to reduce exploitation depending on the length of time since the last mark–recapture population estimate for a population (Hansen et al. 1991). Three years after a mark–recapture population estimate, a lake area–walleye abundance relationship was formerly used to estimate a safe harvest quota in the tribal subsistence fishery, and angler daily bag limits were negatively correlated with tribal harvest (e.g., 5, 3, 2 walleye/angler/day) (Nate et al. 2000; Beard et al. 2003). Angler regulations also included a 381 mm minimum length limit in most cases. Given downward trajectories in walleye natural recruitment and adult abundance, recent adjustments have made to this management system to reduce exploitation rates. Currently, the limit reference point exploitation rate remains 35% of an adult population. Tribal subsistence harvest quotas are now based on a mixed model ≥3 years post a mark–recapture population estimate, and angling regulations generally consist of a 381 mm minimum length limit, 508 – 610 mm protected no harvest slot length limit, and a daily bag limit of three walleye with only one walleye ≥610 mm allowed to be harvested (Hansen et al. 2015b). These changes were made to provide greater harvest opportunity and equity in the tribal subsistence fishery, to reduce recreational angling exploitation rates, and to increase the proportion of longer females in a population. Last, a pulse fishing rule is in place within the tribal subsistence fishery such that tribal quotas alternate between ≤95% and ≤60% of safe harvest between years. This joint management system is heavily reliant upon the assumption of 35% of the adult walleye population being a sustainable, long-term exploitation rate and consistent natural recruitment. If these assumptions are not met, there is a high likelihood for walleye population decline. In other joint walleye fisheries such as those in Minnesota, output controls are also used in the recreational fishery; however, tribal harvest quotas are based on biomass, and in some cases (e.g., Mille Lacs), additional seasonal closures may be enacted on the recreational fishery. Under these current joint walleye management systems, I question what may happen when traditional assumptions of fisheries management do not hold and propose alternatives to manage these joint fisheries in the face of declining natural recruitment and walleye abundances.
Three common assumptions in joint walleye management systems are (1) a stable, sustainable exploitation rate; (2) consistent natural recruitment (i.e., compensation); and (3) that tribal subsistence and recreational angling exploitation influence walleye populations the same way. In the Ceded Territories of Wisconsin, the 35% limit reference point exploitation rate was established from long-term compulsory creel census data from unregulated walleye recreational fishing (i.e., no minimum length limit, no bag limit, no closed season) on Escanaba Lake, Vilas County, Wisconsin (Sass et al. 2022b). During the mid-1950s–mid-1980s, the walleye population in Escanaba Lake was sustainable under an average exploitation rate of 35% in the recreational fishery, and this was used to establish the limit reference point exploitation rate in the Ceded Territories of Wisconsin joint walleye management system. Escanaba Lake was not subject to tribal subsistence exploitation during this period. Escanaba Lake's walleye population was assumed to be representative of all walleye populations in northern Wisconsin. Given this assumption and recent declines in walleye natural recruitment and population abundances, the 35% limit reference point exploitation rate has been questioned and several experimental, empirical, and modeling studies have been conducted to estimate a sustainable exploitation rate representative of most walleye populations.
Based on growing degree days in northern Wisconsin, Lester et al. (2014) predicted that a sustainable annual walleye exploitation rate would be about 20%, although growing degree days in Wisconsin are increasing due to climate change (Hansen et al. 2017). Using walleye production estimates, Rypel et al. (2015) suggested that a 20% exploitation rate equated to average harvest of annual walleye production in Escanaba Lake during 1965–2009. In Rypel et al. (2015), 35% annual exploitation would on average harvest 37% more than annual production, which should theoretically result in a decline in the walleye population. Using a statistical catch-at-age model, Tsehaye et al. (2016) estimated that an exploitation rate of about 20% would be appropriate to meet management goals of adult abundance ≥7.4 adults/ha for a general walleye lake in the Ceded Territories of Wisconsin. In contrast, Schueller et al. (2008) predicted that walleye populations could sustain 60%–85% annual exploitation; however, the unfished equilibrium biomass used in the model was unrealistically high. Experimental exploitation rate studies on Big Crooked Lake, WI (35% annual exploitation; Sass and Shaw 2018) and Sherman Lake, WI (50% annual exploitation; Sass et al. 2022c) suggested that these exploitation rates were sustainable over a 10-year period for these naturally recruiting populations; however, other walleye population demographic changes were observed. In Rypel et al. (2015), Sass and Shaw (2018), and Sass et al. (2022c), the key to long-term sustainability under these exploitation rates was consistent natural recruitment and elevated survivorship of age-0 and age-1 fish. These walleye populations may be the anomaly given long-term natural recruitment declines observed in other midwestern walleye populations. As such, limit reference point exploitation rates should likely be more conservative and representative of most walleye populations (≤20%) (Tsehaye et al. 2016). Further, limit reference point exploitation rates should be consistently and iteratively revisited to make necessary adjustments based on available data and walleye population status and trends. Current Ceded Territories of Wisconsin walleye exploitation rates in the joint fishery rarely exceed 35% and average about 11%–14% per year. Nevertheless, ≥20% is exceeded much more frequently than 35%. My perspective and review suggest that the limit reference point exploitation rate should be ≤20%. And, perhaps even more stringent if walleye natural recruitment continues to decline (Rypel et al. 2018) because depensatory thresholds likely differ among individual walleye populations (Fig. 3).
Rypel et al. (2018) suggested that Ceded Territories of Wisconsin walleye exploitation rates may be consistently exceeding annual production based on significant declines in biomass, production, and production:biomass ratios (i.e., population replacement rate) over time. These findings were commensurate with consistent annual exploitation rates of walleye populations with a diminished capacity to replace biomass through natural recruitment or stocking. Theoretically, such harvest management would result in reduced adult population abundance with declining natural recruitment. An experiment on Escanaba Lake is currently ongoing to test for the effects of 10 years of annual 150% production exploitation to test for walleye population demographic, natural recruitment, and adult abundance responses (Sass et al. 2022b). This experiment includes tribal subsistence harvest based on the current management system and recreational angler exploitation using the current Ceded Territories of Wisconsin general walleye harvest regulations (381 mm minimum length limit, 508–610 mm protected no harvest slot length limit, daily bag limit of three fish with only fish ≥610 mm) (Sass et al. 2022b). Taking this a step further, Embke et al. (2019) and Mrnak et al. (2024) used walleye production and yield:production ratios to confirm that production overharvest was occurring on some Ceded Territories of Wisconsin walleye populations. The findings of Tsehaye et al. (2016), Rypel et al. (2018), Embke et al. (2019), Sass et al. (2021), and Mrnak et al. (2024) suggest and support that walleye lakes have different productive capacities (Ryder 1965), which may necessitate population-specific harvest management for sustainability (Fig. 3). Unfortunately, population-specific management is difficult, given the number of walleye lakes in the midwestern United States and Canada and regulation complexity is unpalatable to many anglers and tribal members. An alternative is to manage all walleye populations in the joint fishery precautionarily by reducing the current limit reference point exploitation rate to ≤20%, with continued fisheries-dependent and -independent monitoring to inform whether further limit reference point exploitation rate reductions are warranted to sustain remaining walleye populations. Another alternative would be to manage joint fisheries resources based on yield:production ratios to ensure that ≤ annual production is harvested. Again, this would not likely be feasible for individual lake walleye management. Moving forward, the most precautionary approach appears to be a reduction in the limit reference point exploitation rate, consistent monitoring, and annual evaluations of yield:production ratios. Collectively, this management approach should serve to reduce walleye exploitation rates and allow for responsive adjustments should walleye natural recruitment, adult population abundance, and productivity continue to decline.
Factors negatively influencing walleye populations
Many compounding and interacting factors have been implicated in walleye natural recruitment and population declines (Fig. 2). These stressors can be categorized as slow-moving environmental or unmanageable (Carpenter et al. 2017), abrupt or increasingly variable, harvest or fisher-related, and ecosystem-based (e.g., fish community, food web). Regardless of the stressors or their interactions, these variables are negatively influencing walleye natural recruitment and challenging the persistence of walleye populations. Walleye are no longer the dominant species in many former populations, which can lead to positive feedback loops being reinforced to suppress walleye populations, increase the probability of crossing a depensatory threshold, and challenge re-establishment of dominance (Sass et al. 2021; Dassow et al. 2023).
Slow-moving environmental or unmanageable drivers (Carpenter et al. 2017) include climate change, lake warming, and loss of suitable thermal–optical habitat for walleye. Climate change (i.e., increases in growing degree days) and lake warming have been consistent predictors implicated in walleye natural recruitment declines (Hansen et al. 2017; Dassow et al. 2022; Feiner et al. 2022a). Increases in growing degree days have also been predicted to favor warmwater centrarchids (e.g., largemouth bass, black crappie, bluegill) over coolwater fishes such as walleye (Hansen et al. 2017, 2018; Broda et al. 2022; Embke et al. 2022; Zebro et al. 2022). The interaction of climate change, lake warming, and centrarchid relative abundances have been linked to a higher probability of depensatory recruitment dynamics in walleye (Sass et al. 2021; Dassow et al. 2023). Loss of suitable thermal–optical habitat for walleye through climate change has also been suggested to negatively influence walleye natural recruitment and population abundance (Lester et al. 2004; Mahlum et al. 2023). Nevertheless, empirical tests of thermal–optical habitat change and influences on walleye populations have been mixed. Minnesota lakes have shown increases and decreases in walleye thermal–optical habitat over time, with potential negative effects suggested for small length classes of walleye (Mahlum et al. 2023). Vasquez (2024) and Michels (2024) did not observe a strong influence of thermal–optical habitat influencing in situ walleye habitat use and growth of age-0 walleye, respectively. Slow environmental variables are often unmanageable in the short term; thus, actions within managerial control are needed to offset or slow negative influences associated with these drivers of walleye population change (Carpenter et al. 2017).
Abrupt or highly variable stressors include lakeshore residential development, land use change, and phenological variability associated with climate change (Fig. 2). Lakeshore residential development and the intensification of the riparian footprint associated with the colonization of lakeshores by humans has negatively influenced lake ecosystems and fish communities in numerous ways. Lakeshore residential development has been associated with the loss of coarse woody habitat (CWH) and allochthonous sources of energy to lakes (e.g., CWH, leaf litter) (Christensen et al. 1996; Francis and Schindler 2006; Marburg et al. 2006). Allochthonous sources of energy dictate fish community productivity in many north-temperate lakes (Pace et al. 2004; Wilkinson et al. 2013; Sass et al. 2019), and walleye biomass, productivity, and production:biomass ratios have declined significantly over time in northern Wisconsin lakes (Rypel et al. 2018). Lakeshore residential development and land use change can also cause a transition from natural allochthonous inputs driving food webs and water quality to those dominated by excess nutrient loading and autochthony. Eutrophication may negatively influence walleye populations through lake warming and loss of suitable dissolved oxygen concentrations. Phenological variability due to climate change in north-temperate lakes (e.g., more variable ice dynamics) has also been shown to negatively influence walleye recruitment (Feiner et al. 2022a; Barta et al. 2024). Higher age-0 walleye recruitment to the first fall has been associated with long-term average ice out dates; deviations from the long-term average have resulted in poorer age-0 year classes. Climate change influences are often studied as linear increases in temperature predicted over time. For walleye, it appears that directional climate change may be less important to walleye natural recruitment compared to the recent high variability observed in ice dynamics (Feiner et al. 2022a; Sass et al. 2022b; Barta et al. 2024). Phenological variability therefore has the potential to lead to a greater frequency of “mismatches” negatively influencing walleye natural recruitment. Although phenological variability is an abrupt change associated with the slow-moving driver of climate change, managers may have some influence on lakeshore residential development and land use change negatively influencing walleye habitat. For example, the ecological footprint of lakeshore residential development could be managed to allow for allocthonous inputs of energy from riparian zones, and watershed management could consider land use change that may negatively influence walleye or cool and coldwater species habitat (Tingley et al. 2019). Overall, a species cannot persist without suitable habitat (Sass et al. 2017).
A common theme in fisheries management is that exploited fisheries can be considered compensatory resources if managed properly. Traditional fisheries management assumes that controlling exploitation rates through input and output controls and compensatory natural recruitment will allow populations to consistently recover and provide harvest opportunities. Traditional assumptions have been challenged in many ways by not accounting for angler effort dynamics, failures of input and output controls to regulate exploitation, declines and failures in natural recruitment, and the reality of depensation (Post 2013). If traditional fisheries management tenets and assumptions held, exploited fisheries would have a low probability of crossing a depensatory threshold and collapsing. Walleye populations have collapsed and are therefore not immune to traditional single-species fisheries management approaches, which have commonly been used to manage the species. Recent identification of walleye production overharvest (Embke et al. 2019) and depensatory recruitment dynamics (Post et al. 2002; Sass et al. 2021; Cahill et al. 2022; Dassow et al. 2023) foreshadow issues in traditional single-species management approaches that remain rigid to changing walleye population productivity and environmental stressors. Exploitation rates have remained consistent or increased over time, yet production has declined because of natural recruitment declines and the inability of stocking to replenish walleye stocks. Production overharvest will reduce adult walleye population abundance, as evidenced in the Ceded Territories of Wisconsin and other walleye fisheries (Post et al. 2002; Hansen et al. 2015; Pedersen et al. 2018; Rypel et al. 2018; Embke et al. 2019; Cahill et al. 2022). In turn, reductions in adult abundance increase the probability of a depensatory threshold being crossed, leading to population collapse (Sass et al. 2021; Dassow et al. 2023) (Fig. 3). And, exploitation is just one of several factors that may be negatively influencing walleye population sustainability in addition to slow-moving or unmanageable, abrupt or highly variable, and (or) ecosystem-based drivers (Fig. 2).
Ecosystem-based stressors such as species interactions and food web dynamics are also negatively influencing walleye populations (Fig. 2). Of particular importance are aquatic invasive species introductions, fish community composition shifts, angler behaviors, and single-species management in multi-species fisheries. These drivers are integrally linked to ecosystem-based fisheries management and human dimensions of fisheries, and appear to be interacting to create conditions that are inhibiting cultivating influences to promote the persistence of walleye.
Invasive species have had profound effects on native fishes and food webs, and walleye are no exception. Introductions of invasive rainbow smelt (Osmerus mordax (Mitchill)) to inland lakes have often resulted in the loss of walleye natural recruitment, with subsequent extirpations of walleye populations in the absence of intervention (Mercado-Silva et al. 2007; Mrnak et al. 2023). Spiny water flea (Bythotrephes logimanus Leydig) and zebra mussels (Dreissena polymorpha (Pallas)) have also changed water quality conditions and food web interactions potentially to the detriment of suitable walleye habitat (Hansen et al. 2019, 2020; Honsey et al. 2020). Prevention of new introductions and invasive species eradication/control efforts (Gaeta et al. 2015; Mrnak et al. 2023) will be paramount to the sustainability of native species, including walleye.
Climate change and angler behaviors appear to be benefiting warmwater species in lieu of north-temperate fish communities being dominated by cold and coolwater species such as walleye. Abundance increases of black crappie, bluegill, bullheads, and largemouth bass have been observed or predicted to be detrimental to walleye populations through potential competition and predation. A negative relationship has been observed between black and white crappie (Pomoxis annularis Rafinesque) relative abundance and age-0 walleye recruitment (Quist et al. 2003; Broda et al. 2022). Extensive bullhead removals on several northern Wisconsin lakes increased walleye natural recruitment and adult abundances (Sikora et al. 2021). Increases in largemouth bass abundances have been related to declines in walleye natural recruitment (Hansen et al. 2015). Predictions of largemouth bass abundance increases with climate change further exacerbate negative interactions between these species (Hansen et al. 2017, 2018), and whole-lake removals of centrarchids may be insufficient to restore walleye natural recruitment in the short term (Embke et al. 2022). Re-establishment of walleye natural recruitment and fish community dominance appear even more daunting given differences in angler behaviors between fish species, namely, the harvest-oriented nature of walleye fisheries and the increasing prevalence of voluntary-release fisheries for the black basses and panfish such as black crappie and bluegill (Sass and Shaw 2020; Sass et al. 2023).
Despite declines in walleye abundance, walleye angling and tribal subsistence fisheries remain harvest-oriented (Gaeta et al. 2013; Hansen et al. 2015). Angling fisheries for co-occurring species like the black basses and muskellunge (Esox masquinongy Mitchill) are now predominantly catch-and-release (Allen et al. 2008; Gaeta et al. 2013; Hansen et al. 2015; Gilbert and Sass 2016; Eslinger et al. 2017; Hessenauer et al. 2018; Shaw et al. 2019; Sass and Shaw 2020). Black crappie and bluegill anglers are also becoming more catch-and-release-oriented (Sass et al. 2023) due to desires of higher population size structure and concerns of high exploitation rates during spring spawning (Rypel 2015). Management agencies have responded by enacting more conservative harvest regulations on panfish populations such as those in Wisconsin. Catch-and-release practices may result in greater abundances of the black basses, muskellunge, and panfish regardless of harvest regulations, which may be a detriment to walleye population dominance and sustainability. This means that species-specific management goals and angler behaviors in fisheries managed for multiple species may be pitting regulations and angler behaviors against the harvest-oriented nature of walleye fisheries necessitating an ecosystem-based fisheries management approach that considers species interactions, food web effects, and angler desires.
Given the popularity of recreational angling walleye fisheries and tribal subsistence needs, one approach to balance diverse angler desires is “buffet-style” management (van Poorten and Camp 2019) in lake-rich regions. Here, ecosystem-based fisheries management would be used to manage the most productive and exploitation resilient systems suited for walleye specifically for walleye in lieu of other desired species (Rypel et al. 2019; Tingley et al. 2019; Dassow et al. 2022). Alternatively, non-walleye lakes would be managed specifically for species best suited for available within-lake habitat (Rypel et al. 2019; Tingley et al. 2019; Dassow et al. 2022). This also acknowledges that former “walleye lakes” may transition to non-walleye lakes under climate change, thus requiring anglers, tribal subsistence fishers, and stakeholder groups to inevitably accept this change (Feiner et al. 2022b; Shultz et al. 2022). Using this framework, regulations would be species-specific, but would fully acknowledge that trying to manage individual species for certain goals within multi-species fisheries would be difficult and not likely beneficial to walleye. Liberal harvest regulations could also be enacted on alternative species in walleye lakes, although social norms of voluntary catch-and-release may render these liberal regulations ineffective without a change in angler behavior and increased harvest rates (Miranda et al. 2017; Hessnauer et al. 2018; Sass and Shaw 2020).
Depensation
Depensation or positive density dependence is reduced juvenile survival at low adult stock sizes and can be caused by several factors (e.g., fish behavior, angler behavior, management initiatives, food web alterations) (Ricker 1954, 1963; Liermann and Hilborn 1997, 2001; Walters and Kitchell 2001; Post et al. 2002; Hilborn et al. 2014; Sass et al. 2021; Dassow et al. 2023). Depensation can also extend to reduced juvenile survival rates at low age-0 stock size (Zebro et al. 2022). Depensation (i.e., crossing an ecological tipping point or positive biomass threshold) can result in population collapse or slow recovery after the reduction or elimination of harvest due to proposed mechanisms, including (1) reduced probability of fertilization; (2) impaired group dynamics; and (3) conditioning of the environment (i.e., cultivation effects) (Liermann and Hilborn 2001; Walters and Kitchell 2001; Sass et al. 2021; Dassow et al. 2023). Depensation contrasts the long-held theoretical ecological population growth and compensation paradigms (i.e., higher juvenile survivorship at low adult stock sizes or negative density dependence) of populations at low stock sizes. Depensation was anecdotally implicated for the “invisible collapse” of Alberta, Canada walleye fisheries (Post et al. 2002; Cahill et al. 2022) and empirically observed for northern Wisconsin walleye populations (Sass et al. 2021; Dassow et al. 2023) (Fig. 4). For Alberta walleye, the “invisible collapse” was attributed to high angler effort on limited walleye population resources (Post et al. 2002, 2008; Post 2013). In extension, hyperstability in catch rates and (or) anglers not leaving a fishery despite reduced catch rates were likely responsible.
In Wisconsin, depensatory recruitment dynamics and population declines have been attributed to climate change (Hansen et al. 2017, 2018), production overharvest (Embke et al. 2019), invasive species (Mercado-Silva et al. 2007), habitat loss (Sass et al. 2017), angler behavior and single-species management (Hansen et al. 2015; Sass and Shaw 2020), changes in fish community composition (Hansen et al. 2017, 2018; Sikora et al. 2021; Broda et al. 2022; Embke et al. 2022), and their interactions (Dassow et al. 2023). For Wisconsin walleye, these factors are likely leading to depensation due to a loss of cultivation influences of walleye populations at higher abundances (Dassow et al. 2023). After the “invisible collapse” in Alberta, some walleye fisheries recovered over time following fishery closure and stringent regulations, although some populations did not (Cahill et al. 2022). Conservative harvest regulations were rapidly implemented once the issue was identified, and stocking was not a component of rehabilitation efforts in Alberta. Despite clear evidence of walleye natural recruitment and population declines in Wisconsin, self-sustaining populations have continued to be lost over time. Current Wisconsin walleye rehabilitation efforts using conservative harvest regulations, compromise between anglers and tribal subsistence fishers, and stocking have largely been unsuccessful in re-establishing natural recruitment (Embke et al. in review). Alberta walleye fisheries likely recovered and avoided depensation in some cases due to a rapid management response once the issue was identified, low species diversity fisheries, availability of suitable habitat, the lack of reliance on stocking, reduced exploitation, and adult stock sizes not dipping below a critical depensatory threshold. Wisconsin walleye rehabilitations may be more challenging due to competing interests of stakeholders of walleye fishery resources, lakeshore residential development and land use change influences on within-lake habitat, greater fish species diversity, lack of food web effects acknowledgement in multi-species fisheries primarily managed with single-species management, and insufficient reductions in angler and tribal subsistence exploitation.
Despite walleye population declines, walleye angling popularity remains high and tribal subsistence needs have become greater over time. Although equity has been a major focus of management efforts in joint walleye fisheries, more conservative harvest regulations in the angling fishery and small reductions in exploitation rates have been countered by increasing tribal subsistence needs. Therefore, total exploitation in the joint fishery has not been reduced or offset despite declining walleye resources. This can result in production overharvest, where harvest exceeds annual production, which is further exacerbated by natural recruitment declines (Embke et al. 2019). Stocking has been ineffective for completely filling this natural recruitment void (Elwer et al. 2023). Collectively, these dynamics, policies, and desires set the stage for an increased probability of depensatory recruitment dynamics. Even more troubling is the fact that adult stock size critical depensatory thresholds likely differ among walleye populations (Fig. 3). Therefore, the only responsible, precautionary, and logical pathway to prevent or reduce the probability of depensation is to drastically reduce walleye exploitation rates (Rypel et al. 2015; Tsehaye et al. 2016; Carpenter et al. 2017; Embke et al. 2019; Mrnak et al. 2024). That said, this requires that natural recruitment declines are wholly, partially, or interactively associated with high exploitation rates. Unfortunately, it may not be as simple because other drivers are negatively influencing walleye populations, delaying recovery, and working against walleye due to environmental change. Preventing continued walleye declines, and avoiding depensatory thresholds likely requires identifying limiting factor(s) leading to natural recruitment issues and setting the ecological stage to create cultivating conditions for walleye prior to injecting management actions. Cultivating conditions to favor walleye should be considered if they are the species most desired by anglers and tribal subsistence fishers with the understanding that other desirable fish species may not benefit under an ecosystem-based fisheries management framework (Radinger et al. 2023). Although many factors negatively influencing walleye are out of managerial control (Carpenter et al. 2017), those in managerial control such as harvest regulations, habitat conservation and enhancement, some aspects of angler effort, and stocking are. Even though adult stock size has been a poor predictor of walleye recruitment (Madenjian et al. 1996; Hansen et al. 1998; Beard et al. 2003; Tsehaye et al. 2016; Shaw et al. 2018; Feiner et al. 2019; Krabbenhoft et al. 2023), zero adults equal zero recruitment. Thus, conservation of walleye populations through reduced exploitation is important and stocking may not lead to favorable outcomes or recovery for many reasons. Here, ecosystem-based fisheries management may be critical compared to the management panacea of stocking (Arlinghaus et al. 2022; Radinger et al. 2023). Although limited empirical evidence exists for depensation (but see Sass et al. 2021), it is a mechanism by which species or populations go extinct or become extirpated. Although compensation is not infinite, and overharvest can cause a fish population to collapse without depensation, depensatory processes create a positive biomass threshold below which the production–harvest dynamics must result in extirpation (Fig. 1).
Moving forward—avoiding an ecological tipping point
Fisher effort dynamics
Angler and subsistence fisher effort dynamics may dictate movement about a fisheries-rich landscape when catch rates and fisher satisfaction decline with declining fish stocks. Theoretically, this may result in population recovery after fishing and exploitation is reduced. More realistically, average or poor fisheries across a fisheries landscape may manifest if effort in open access fisheries remains high and fishing opportunities are limited (Post et al. 2002, 2008; Wilson et al. 2020). Despite abundant lakes and walleye fisheries in the midwestern United States and Canada, most fisheries are open access, and many are subject to tribal subsistence harvest. Further, anglersheds for this region can be broad (Lant et al. 2022). Walleye catch and harvest rates in angling and tribal fisheries have been shown to be hyperstable (Hansen et al. 2005; Mrnak et al. 2018; Mosley et al. 2022), which may also create a disjunct in angler effort responses and walleye population status. Ultimately, high angler effort, tribal subsistence needs, abiotic and biotic stressors, the life history of walleye, and the harvest-oriented nature of walleye fisheries may result in these fisheries being prone to depensation (Post 2013). Angler and tribal subsistence fisher effort dynamics may push walleye resources below a depensatory threshold such that populations are systematically depleted or slow to recover over time and across a fisheries landscape in the absence of precautionary or proactive management intervention and effort limitations (Post 2013; Sass et al. 2021; Cahill et al. 2022; Dassow et al. 2023). An examination of walleye angler and tribal subsistence effort trends over time during this period of declining abundance would be a fruitful area of research to inform questions about effort dynamics in the future. For example, consistent or increasing directed effort for walleye over time may suggest the need for more conservative input controls, whereas a decline in directed walleye effort may lead to reduced exploitation rates and remaining population resiliency.
Bright spots
The identification of “bright spots” (Bennett et al. 2016) (i.e., walleye populations that have been sustainable in the long term and are predicted to remain resilient to crossing a depensatory threshold) are becoming more common given exploitation and environmental stressors to promote conservation. Walleye “bright spot” populations have generally been associated with high-quality habitat (e.g., low lakeshore residential development, abundant spawning habitat), lake characteristics (e.g., larger surface area, colder water temperatures, lotic influences or characteristics), and food web and fish community characteristics cultivating the persistence of walleye (e.g., presence of yellow perch (Perca flavescens (Mitchill)), low abundances of centrarchids) (Hansen et al. 2015a, 2017; Raabe et al. 2020; Embke et al. 2022; Dassow et al. 2023; Krabbenhoft et al. 2023; Shaw et al. 2023). “Bright spot” populations have also been related to walleye population productivity and lower exploitation rates relative to production (Rypel et al. 2018; Embke et al. 2019; Mrnak et al. 2024). “Bright spot” walleye populations have been less related to specialized walleye harvest regulations. Although this concept appears to make ecological sense for conservation, “bright spot” identification can be detrimental as these populations may also draw increased attention and effort. For example, clean water lakes may draw more recreational activities and lakeshore residential development (Garrah et al. 2019). Similarly, “bright spots” in depleted fisheries landscapes are likely to draw more effort and exploitation. In these cases, “bright spots” should be protected the most conservatively with particular attention to fisheries-dependent and -independent monitoring and alternatives to traditional input and output controls.
Several methods and an example to avoid the “tragedy of the bright spots” include the conservation of coldwater fisheries resources in Minnesota. Here, “bright spots” supporting native cisco (Coregonus artedi Lesueur), lake whitefish (Coregonus clupeaformis (Mitchill)), and lake trout (Salvelinus namaycush (Walbaum)) have been identified and a watershed scale conservation approach has been used to prevent land use change to maintain coldwater resources and fisheries through dedicated funding (Tingley et al. 2019). An analogous approach could be used for walleye, considering habitat needs and reduced exploitation. Likewise, decision support tools, which consider future predictions based on environmental stressors could be used to identify “bright spots” for conservation as well as walleye population trajectories not likely to change despite management intervention (Tingley et al. 2019; Dassow et al. 2022). In the case of walleye “bright spots”, identification should be used for aggressive and proactive conservation and not to draw attention to remaining harvest opportunities for more effort and exploitation.
Concluding remarks
My review and perspective of walleye population declines suggested that exploited fisheries should be viewed and managed in the context of resiliency and aversion to crossing an ecological tipping point or depensatory threshold. In fact, much of the discussion provided herein provides empirical support for the conceptual arguments of Post (2013). When drivers outside of managerial control are causing fish population declines, it is logical to consider exploited fisheries in the context of resilience to depensation and thus requiring management interventions within managerial control such as reductions in exploitation to sustain populations. However, interventions within managerial control may be rendered ineffective when exploitation is not sufficiently reduced, angler desires and tribal subsistence needs trump conservation and sustainability, traditional single-species fisheries management in fisheries managed for multiple species is the norm, angler behaviors and regulations pit species outcomes and management goals against one another, and effort dynamics of anglers and tribal subsistence fishers behave differently than assumed. In these cases, traditional fisheries management approaches (single-species management) and assumptions of exploited fisheries (e.g., compensation, fishers leaving fisheries when catch rates decline) may not hold and fisheries have the potential to collapse. Overfishing and depensation are real. Exploited populations that are also subject to various stressors and disturbances would not collapse, be extirpated, or go extinct if this were not the case. For walleye, it appears that immediate action is needed to conserve remaining populations to avoid their systematic depletion. Currently, it appears as if naturally recruiting walleye populations are being depleted along an angler and tribal subsistence effort front, with stocking primarily being used as a last ditch effort for species persistence even when a depensatory threshold may have already been crossed (Sass et al. 2021; Dassow et al. 2023) (Fig. 4). At best, stocking is supporting low quality, put–grow–take fisheries with little evidence of re-establishing self-sustaining populations and meeting angler desires and tribal subsistence needs. If this continues, many walleye populations are unlikely to exist in the short and long term due to environmental stressors, food web change preventing recovery, and depensatory recruitment dynamics. Stocking will not be able to meet the needs of tribal subsistence fishers, nor angler desires. Anglers are mobile and specialized, which may suggest that specialized walleye anglers and tribal subsistence fishers will continue to range farther to exploit remaining harvest opportunities, leaving stocked fisheries behind. My review and perspective suggest that walleye exploitation rates be immediately reduced to be protective of remaining populations and to allow for greater opportunity for multiple spawning events and a higher probability of recruitment “matches”, given current stressors negatively influencing walleye populations and natural recruitment. Immediate reductions in walleye exploitation may also buy time for the identification of limiting factors to natural recruitment that allow for management interventions to set the ecological stage to cultivate conditions most favorable to walleye and long-term persistence.
My review and perspective also clearly showed that traditional fisheries management tenets (e.g., compensation, single-species management) are being challenged in complex, exploited fisheries, such as those for walleye. Fisheries management systems are complex and includes aspects of ecology, fish, habitat, and human dimensions (Sass and Allen 2014). As such, ecosystem-based fisheries management and recognition of fisheries as social–ecological systems may be more important than ever for sustaining exploited fisheries. Although some progress has been made on both fronts, additional research and successful examples are needed to garner angler, tribal subsistence fisher, and stakeholder support to fully transition to such approaches as the norm in fisheries management. Radinger et al. (2023) provided an excellent example and test of the influences of ecosystem-based fisheries management compared to single-species management (i.e., stocking) on fish production. Ecosystem-based fisheries management unequivocally outperformed single-species management. This will also require a concerted effort to train the next generation of fisheries students, managers, researchers, and scientists in ecology, ecosystem science, ecosystem-based fisheries management, and the human dimensions of fisheries. If the legacy effects of single-species fisheries management persist, it is likely that other exploited fisheries will cross ecological tipping points like that of many walleye populations.
Acknowledgements
I would like to thank the Wisconsin Department of Natural Resources (WDNR) walleye species team, the Great Lakes Indian Fish and Wildlife Commission (GLIFWC), and the Walleye Technical Working Group comprised of WDNR fisheries biologists and researchers, GLIFWC fisheries biologists and scientists, and Ojibwe tribal representatives for ongoing discussions leading to this review and perspective. Special thanks to Zach Feiner for developing the map shown in Fig. 4. I would also like to thank Dr. John Post and an anonymous reviewer for providing constructive reviews that greatly improved this manuscript. This research was supported by the U.S. Fish and Wildlife Service Federal Aid in Sportfish Restoration program and the WDNR.
References
Allen M.S., Walters C.J., Myers R. 2008. Temporal trends in largemouth bass mortality, with fishery implications. North American Journal of Fisheries Management, 28(2): 418–427.
Arlinghaus R., Riepe C., Theis S., Pagel T., Fujitani M. 2022. Dysfunctional information feedbacks cause the emergence of management panaceas in social–ecological systems: the case of fish stocking in inland recreational fisheries. Journal of Outdoor Recreation and Tourism, 38: 100475.
Bailey C.T., Noring A.M., Shaw S.L., Sass G.G. 2019. Live versus artificial bait influences on walleye (Sander vitreus) angler effort and catch rates on Escanaba Lake, Wisconsin, 1993–2015. Fisheries Research, 219: 105330.
Barta M.E., Sass G.G., Reed J.R., Cichosz T.A, Shultz A.D., Luehring M., Feiner Z.S. 2024. Lagging spawning and increasing phenological extremes jeopardize walleye (Sander vitreus) in north temperate lakes. Limnology and Oceanography Letters 9: 229–236.
Barton B.A. 2011. Biology, management, and culture of walleye and sauger. American Fisheries Society, Bethesda, Maryland.
Beard T.D. Jr., Hansen M.J., Carpenter S.R. 2003. Development of a regional stock–recruitment model for understanding factors affecting walleye recruitment in northern Wisconsin lakes. Transactions of the American Fisheries Society, 132: 382–391.
Bellido J.M., Sumaila U.R., Sánchez-Lizaso J.L., Palomares M.L., Pauly D. 2020. Input versus output controls as instruments for fisheries management with a focus on Mediterranean fisheries. Marine Policy, 118: 103786.
Bennett E.M., Solan M., Biggs R., McPhearson T., Norström A.V., Olsson P., et al. 2016. Bright spots: seeds of a good Anthropocene. Frontiers in Ecology and the Environment, 14: 441–448.
Boehm H.I. 2016. Identifying recruitment bottlenecks for age-0 walleye Sander vitreus in northern Wisconsin lakes. M.S. Thesis, University of Wisconsin-Stevens Point, College of Natural Resources.
Bootsma M.L., Miller L., Sass G.G., Euclide P.T., Larson W.A. 2021. The ghosts of propagation past: haplotype information clarifies the relative influence of stocking history and phylogeographic processes on contemporary population structure of walleye (Sander vitreus). Evolutionary Applications, 14(4): 1124–1144.
Boxall P.C. 1995. The economic value of lottery-rationed recreational hunting. Canadian Journal of Agricultural Economics/Revue canadienne d'agroeconomie, 43: 119–131.
Broda S.P., Feiner Z.S., Mrnak J.T., Shaw S.L., Sass G.G. 2022. Black crappie influences on walleye natural recruitment in northern Wisconsin lakes. North American Journal of Fisheries Management, 42: 1202–1214.
Bruch R.M. 1999. Management of lake sturgeon on the Winnebago System—long term impacts of harvest and regulations on population structure. Journal of Applied Ichthyology, 15: 142–152.
Cahill C.L., Walters C.J., Paul A.J., Sullivan M.G., Post J.R. 2022. Unveiling the recovery dynamics of walleye after the invisible collapse. Canadian Journal of Fisheries and Aquatic Sciences, 79: 708–723.
Carpenter S.R., Brock W.A., Hansen G.J., Hansen J.F., Hennessy J.M., Isermann D.A., et al. 2017. Defining a safe operating space for inland recreational fisheries. Fish and Fisheries, 18(6): 1150–1160.
Christensen D.L., Herwig B.R., Schindler D.E., Carpenter S.R. 1996. Impacts of lakeshore residential development on coarse woody debris in north temperate lakes. Ecological Applications, 6(4): 1143–1149.
Cooke S.J., Venturelli P., Twardek W.M., Lennox R.J., Brownscombe J.W., Skov C., et al. 2021. Technological innovations in the recreational fishing sector: implications for fisheries management and policy. Reviews in Fish Biology and Fisheries, 31: 253–288.
Cox S.P., Beard T.D., Walters C. 2002. Harvest control in open-access sport fisheries: hot rod or asleep at the reel? Bulletin of Marine Science, 70: 749–761.
Cushing D.H. 1990. Plankton production and year-class strength in fish populations: an update of the match/mismatch hypothesis. In Advances in marine biology. Vol. 26. Academic Press. pp. 249–293.
Dassow C.J., Ross A.J., Jensen O.P., Sass G.G., van Poorten B.T., Solomon C.T., Jones S.E. 2020. Experimental demonstration of catch hyperstability from habitat aggregation, not effort sorting, in a recreational fishery. Canadian Journal of Fisheries and Aquatic Sciences, 77(4): 762–769.
Dassow C.J., Latzka A.W., Lynch A.J., Sass G.G., Tingley R.W. III, Paukert C.P. 2022. A resist-accept-direct decision-support tool for walleye Sander vitreus (Mitchill) management in Wisconsin. Fisheries Management and Ecology, 29(4): 378–391.
Dassow C., Sass G., Shaw S., Feiner Z., Nieman C., Jones S. 2023. Depensation in fish recruitment driven by context-dependent interactions with another predator. Fisheries Research, 262: 106675.
Elwer B.M., VanDeHey J.A., Shaw S.L., Sikora L.W., Mrnak J.T., Sass G.G. 2023. Evaluation of survival and cost to harvestable age of stocked fall fingerling walleye (Sander vitreus) in Northern Wisconsin Lakes. Fisheries Research, 266: 106758.
Embke H.S., Rypel A.L., Carpenter S.R., Sass G.G., Ogle D., Cichosz T., et al. 2019. Production dynamics reveal hidden overharvest of inland recreational fisheries. Proceedings of the National Academy of Sciences, 116: 24676–24681.
Embke H.S., Carpenter S.R., Isermann D.A., Coppola G., Beard T.D. Jr, Lynch A.J., et al. 2022. Resisting ecosystem transformation through an intensive whole-lake fish removal experiment. Fisheries Management and Ecology, 29: 364–377.
Embke H.S., Feiner Z.S., Hansen G.J.A., Mrnak J.T., Rounds C., Sass G.G., et al. Healing ogaa (walleye Sander vitreus) waters: lessons and future directions for inland fisheries rehabilitation. Reviews in Fisheries Science and Aquaculture, in review.
Eslinger L.D., Sass G.G., Shaw S.L., Newman S.P. 2017. Muskellunge population responses to angler catch-and-release practices in Escanaba Lake, Wisconsin, 1987–2014. In Muskellunge management: fifty years of cooperation among anglers, scientists and fisheries biologists. Edited by K.L. Kapuscinski, T.D. Simonson, D.P. Crane, S.J. Kerr, J.S. Diana, J.M. Farrell. American Fisheries Society, Symposium 85, Bethesda, Maryland. pp. 337–355.
Feiner Z.S., Shaw S.L., Sass G.G. 2019. Influences of female body condition on recruitment success of walleye (Sander vitreus) in Wisconsin lakes. Canadian Journal of Fisheries and Aquatic Sciences, 76: 2131–2144.
Feiner Z.S., Wolter M.H., Latzka A.W. 2020a. “I will look for you, I will find you, and I will [harvest] you”: persistent hyperstability in Wisconsin's recreational fishery. Fisheries Research, 230: 105679.
Feiner Z.S., Latzka A.W., Wolter M.H., Eslinger L.D., Hatzenbeler G.R. 2020b. Assessing the rage against the machines: do ice anglers’ electronics improve catch and harvest rates? Fisheries, 45(6): 327–333.
Feiner Z.S., Dugan H.A., Lottig N.R., Sass G.G., Gerrish G.A. 2022a. A perspective on the ecological and evolutionary consequences of phenological variability in lake ice on north-temperate lakes.Canadian Journal of Fisheries and Aquatic Sciences, 79: 1590–1604.
Feiner Z.S., Shultz A.D., Sass G.G., Trudeau A., Mitro M.G., Dassow C.J., et al. 2022b. Resist-accept-direct (RAD) considerations for climate change adaptation in fisheries: the Wisconsin experience. Fisheries Management and Ecology, 29(4): 346–363.
Francis T.B., Schindler D.E. 2006. Degradation of littoral habitats by residential development: woody debris in lakes of the Pacific Northwest and Midwest, United States. AMBIO: A Journal of the Human Environment, 35(6): 274–280.
Gaeta J.W., Beardmore B., Latzka A.W., Provencher B., Carpenter S.R. 2013. Catch-and-release rates of sport fishes in northern Wisconsin from an angler diary survey. North American Journal of Fisheries Management, 33: 606–614.
Gaeta J.W., Hrabik T.R., Sass G.G., Roth B.M., Gilbert S.J., Vander Zanden M.J. 2015. A whole-lake experiment to control invasive rainbow smelt (Actinopterygii, Osmeridae) via overharvest and a food web manipulation. Hydrobiologia, 746: 433–444.
Gilbert S.J., Sass G.G. 2016. Trends in a northern Wisconsin muskellunge fishery: results from a countywide angling contest, 1964–2010. Fisheries Management and Ecology, 23(2): 172–176.
Garrah J., Frei B., Bennett E.M. 2019. Bright spots among lakes in the Rideau Valley Watershed, Ontario. Ecology and Society, 24(3).
Gostiaux J., Boehm H.I., Jaksha N.J., Dembkowski D.J., Hennessy J.M., Isermann D.A. 2022. Recruitment bottlenecks for age-0 Walleye in northern Wisconsin lakes. North American Journal of Fisheries Management, 42: 507–522.
Grausgruber E.E., Weber M.J. 2021a. Shift happens: evaluating the ability of autumn stocked walleye Sander vitreus to shift to natural prey. Fisheries Management and Ecology, 28: 516–527.
Grausgruber E.E., Weber M.J. 2021b. Effects of stocking transport duration on age 0 Walleye. Journal of Fish and Wildlife Management, 12: 70–82.
Haglund J.M., Isermann D.A., Sass G.G. 2016. Walleye population and fishery responses after elimination of legal harvest on Escanaba Lake, Wisconsin. North American Journal of Fisheries Management, 36(6): 1315–1324.
Hansen G.J., Carpenter S.R., Gaeta J.W., Hennessy J.M., Vander Zanden M.J. 2015a. Predicting walleye recruitment as a tool for prioritizing management actions. Canadian Journal of Fisheries and Aquatic Sciences, 72(5): 661–672.
Hansen G.J., Hennessy J.M., Cichosz T.A., Hewett S.W. 2015b. Improved models for predicting walleye abundance and setting safe harvest quotas in northern Wisconsin lakes. North American Journal of Fisheries Management, 35(6): 1263–1277.
Hansen G.J., Read J.S., Hansen J.F., Winslow L.A. 2017. Projected shifts in fish species dominance in Wisconsin lakes under climate change. Global Change Biology, 23: 1463–1476.
Hansen G.J., Midway S.R., Wagner T. 2018. Walleye recruitment success is less resilient to warming water temperatures in lakes with abundant largemouth bass populations. Canadian Journal of Fisheries and Aquatic Sciences, 75: 106–115.
Hansen G.J., Winslow L.A., Read J.S., Treml M., Schmalz P.J., Carpenter S.R. 2019. Water clarity and temperature effects on walleye safe harvest: an empirical test of the safe operating space concept. Ecosphere, 10(5): e02737.
Hansen G.J., Ahrenstorff T.D., Bethke B.J., Dumke J.D., Hirsch J., Kovalenko K.E., et al. 2020. Walleye growth declines following zebra mussel and Bythotrephes invasion. Biological Invasions, 22(4): 1481–1495.
Hansen J.F., Sass G.G., Gaeta J.W., Hansen G.J.A., Isermann D.A., Lyons J., Vander Zanden M.J. 2015. Largemouth Bass management in Wisconsin: intraspecific and interspecific implications of abundance increases. In American Fisheries Society Symposium. Vol. 82. pp. 193–206.
Hansen M.J., Staggs M.D., Hoff M.H. 1991. Derivation of safety factors for setting harvest quotas on adult walleyes from past estimates of abundance. Transactions of the American Fisheries Society, 120(5): 620–628.
Hansen M.J., Bozek M.A., Newby J.R., Newman S.P., Staggs M.D. 1998. Factors affecting recruitment of walleyes in Escanaba Lake, Wisconsin, 1958–1996. North American Journal of Fisheries Management, 18: 764–774.
Hansen M.J., Beard T.D. Jr, Hewett S.W. 2005. Effect of measurement error on tests of density dependence of catchability for walleyes in northern Wisconsin angling and spearing fisheries. North American Journal of Fisheries Management, 25: 1010–1015.
Henderson B.A., Collins N., Morgan G.E., Vaillancourt A. 2003. Sexual size dimorphism of walleye (Stizostedion vitreum vitreum). Canadian Journal of Fisheries and Aquatic Sciences, 60: 1345–1352.
Hessenauer J.M., Vokoun J., Davis J., Jacobs R., O'Donnell E. 2018. Size structure suppression and obsolete length regulations in recreational fisheries dominated by catch-and-release. Fisheries Research, 200: 33–42.
Hilborn R., Hively D.J., Jensen O.P., Branch T.A. 2014. The dynamics of fish populations at low abundance and prospects for rebuilding and recovery. ICES Journal of Marine Science, 71(8): 2141–2151.
Honsey A.E., Feiner Z.S., Hansen G.J. 2020. Drivers of walleye recruitment in Minnesota's large lakes. Canadian Journal of Fisheries and Aquatic Sciences, 77(12): 1921–1933.
Kitchell J.F., Johnson M.G., Minns C.K., Loftus K.H., Greig L., Olver C.H. 1977. Percid habitat: the river analogy. Journal of the Fisheries Research Board of Canada, 34: 1936–1940.
Krabbenhoft C.A., Ludsin S.A., Marschall E.A., Budnik R.R., Almeida L.Z., Cahill C.L., et al. 2023. Synthesizing professional opinion and published science to build a conceptual model of Walleye recruitment. Fisheries, 48(4): 141–156.
Lant M.J., Ogle D.H., Feiner Z.S., Sass G.G. 2022. A comparison of fish catch rates among local, non-local, and non-resident anglers of three northern Wisconsin lakes. Fisheries Research, 250: 106286.
Lawson Z.J., Latzka A.W., Eslinger L. 2022. Stocking practices and lake characteristics influence probability of stocked walleye survival in Wisconsin's Ceded Territory Lakes. North American Journal of Fisheries Management, 42: 523–534.
Lester N.P., Dextrase A.J., Kushneriuk R.S., Rawson M.R., Ryan P.A. 2004. Light and temperature: key factors affecting walleye abundance and production. Transactions of the American Fisheries Society, 133: 588–605.
Lester N.P., Shuter B.J., Venturelli P., Nadeau D. 2014. Life-history plasticity and sustainable exploitation: a theory of growth compensation applied to walleye management. Ecological Applications, 24(1): 38–54.
Li J., Cohen Y., Schupp D.H., Adelman I.R. 1996. Effects of walleye stocking on population abundance and fish size. North American Journal of Fisheries Management, 16(4): 830–839.
Liermann M., Hilborn R. 1997. Depensation in fish stocks: a hierarchic Bayesian meta-analysis. Canadian Journal of Fisheries and Aquatic Sciences, 54(9): 1976–1984.
Liermann M., Hilborn R. 2001. Depensation: evidence, models and implications. Fish and Fisheries, 2(1): 33–58.
Lorenzen K. 2014. Managing fisheries enhancements. In Foundations of fisheries science. Edited by G.G. Sass, M.S. Allen. American Fisheries Society, Bethesda, Maryland. pp. 649–793.
Madenjian C.P., Tyson J.T., Knight R.L., Kershner M.W., Hansen M.J. 1996. First-year growth, recruitment, and maturity of walleyes in western Lake Erie. Transactions of the American Fisheries Society, 125(6): 821–830.
Mahlum S., Vitense K., Corson-Dosch H., Platt L., Read J.S., Schmalz P.J., et al. 2023. Connecting habitat to species abundance: the role of light and temperature on the abundance of walleye in lakes. Canadian Journal of Fisheries and Aquatic Sciences, 80: 273–286.
Marburg A.E., Turner M.G., Kratz T.K. 2006. Natural and anthropogenic variation in coarse wood among and within lakes. Journal of Ecology, 94: 558–568.
Mercado-Silva N., Sass G.G., Roth B.M., Gilbert S.J., Vander Zander M.J. 2007. Walleye recruitment decline as a consequence of rainbow smelt invasions in Wisconsin lakes. Canadian Journal of Fisheries and Aquatic Sciences, 64: 1543–1550.
Michels N.O. 2024. Prey behavior to predator growth: the influence of light intensity on predator–prey interactions in north-temperate lakes and rivers. Ph.D. dissertation, University of Minnesota-Duluth.
Miranda L.E., Colvin M.E., Shamaskin A.C., Bull L.A., Holman T., Jones R. 2017. Length limits fail to restructure a largemouth bass population: a 28-year case history. North American Journal of Fisheries Management, 37(3): 624–632.
Mosley C.L., Dassow C.J., Caffarelli J., Ross A.J., Sass G.G., Shaw S.L., et al. 2022. Species differences, but not habitat, influence catch rate hyperstability across a recreational fishery landscape. Fisheries Research, 255: 106438.
Mrnak J.T., Shaw S.L., Eslinger L.D., Cichosz T.A., Sass G.G. 2018. Characterizing the angling and tribal spearing walleye fisheries in the Ceded Territory of Wisconsin, 1990–2015. North American Journal of Fisheries Management, 38: 1381–1393.
Mrnak J.T., Sikora L.W., Zanden M.J.V., Sass G.G. 2023. Applying panarchy theory to aquatic invasive species management: a case study on invasive rainbow smelt Osmerus mordax. Reviews in Fisheries Science & Aquaculture, 31(1): 66–85.
Mrnak J.T., Embke H.S., Wilkinson M.V., Shaw S.L., Vander Zanden M.J., Sass G.G. 2024. Diminishing productivity and hyperstable harvest in northern Wisconsin walleye fisheries. Canadian Journal of Fisheries and Aquatic Sciences, 81: 1650–1665, (ja).
Nate N.A., Bozek M.A., Hansen M.J., Hewett S.W. 2000. Variation in walleye abundance with lake size and recruitment source. North American Journal of Fisheries Management, 20(1): 119–126.
Neely B.C., Koch J.D., Gido K.B. 2023. Evaluating effects of live-imaging sonar on angler catch of crappies in a Kansas impoundment. Fisheries, 48(2): 49–53.
Pace M.L., Cole J.J., Carpenter S.R., Kitchell J.F., Hodgson J.R., Van de Bogert M.C., et al. 2004. Whole-lake carbon-13 additions reveal terrestrial support of aquatic food webs. Nature, 427(6971): 240–243.
Pedersen E.J., Goto D., Gaeta J.W., Hansen G.J., Sass G.G., Vander Zanden M.J., et al. 2018. Long-term growth trends in northern Wisconsin walleye populations under changing biotic and abiotic conditions. Canadian Journal of Fisheries and Aquatic Sciences, 75: 733–745.
Post J.R. 2013. Resilient recreational fisheries or prone to collapse? A decade of research on the science and management of recreational fisheries. Fisheries Management and Ecology, 20: 99–110.
Post J.R., Sullivan M., Cox S., Lester N.P., Walters C.J., Parkinson E.A., et al. 2002. Canada's recreational fisheries: the invisible collapse? Fisheries, 27(1): 6–17.
Post J.R., Persson L., Parkinson E.V., Kooten T.V. 2008. Angler numerical response across landscapes and the collapse of freshwater fisheries. Ecological Applications, 18(4): 1038–1049.
Quist M.C., Guy C.S., Stephen J.L. 2003. Recruitment dynamics of walleyes (Stizostedion vitreum) in Kansas reservoirs: generalities with natural systems and effects of a centrarchid predator. Canadian Journal of Fisheries and Aquatic Sciences, 60: 830–839.
Raabe J.K., VanDeHey J.A., Zentner D.L., Cross T.K., Sass G.G. 2020. Walleye inland lake habitat: considerations for successful natural recruitment and stocking in North Central North America. Lake and Reservoir Management, 36: 335–359.
Radinger J., Matern S., Klefoth T., Wolter C., Feldhege F., Monk C.T., Arlinghaus R. 2023. Ecosystem-based management outperforms species-focused stocking for enhancing fish populations. Science, 379(6635): 946–951.
Radomski P.J. 2022. Walleye: a beautiful fish of the dark. University of Minnesota Press, Minneapolis, Minnesota.
Rennie M.D., Purchase C.F., Lester N., Collins N.C., Shuter B.J., Abrams P.A. 2008. Lazy males? Bioenergetic differences in energy acquisition and metabolism help to explain sexual size dimorphism in percids. Journal of Animal Ecology, 77: 916–926.
Ricker W.E. 1954. Stock and recruitment. Journal of the Fisheries Research Board of Canada, 11(5): 559–623.
Ricker W.E. 1963. Big effects from small causes: two examples from fish population dynamics. Journal of the Fisheries Research Board of Canada, 20(2): 257–264.
Ricker W.E. 1975. Computation and interpretation of biological statistics of fish populations. Bulletin of the Fisheries Research Board of Canada, 191: 1–382.
Ryder R.A. 1965. A method for estimating the potential fish production of north-temperate lakes. Transactions of the American Fisheries Society, 94: 214–218.
Rypel A.L. 2015. Effects of a reduced daily bag limit on bluegill size structure in Wisconsin lakes. North American Journal of Fisheries Management, 35: 388–397.
Rypel A.L., Goto D., Sass G.G., Vander Zanden M.J. 2015. Production rates of walleye and their relationship to exploitation in Escanaba Lake, Wisconsin, 1965–2009. Canadian Journal of Fisheries and Aquatic Sciences, 72: 834–844.
Rypel A.L., Goto D., Sass G.G., Vander Zanden M.J. 2018. Eroding productivity of walleye populations in northern Wisconsin lakes. Canadian Journal of Fisheries and Aquatic Sciences, 75: 2291–2301.
Rypel A.L., Simonson T.D., Oele D.L., Griffin J.D., Parks T.P., Seibel D., et al. 2019. Flexible classification of lakes for improved fisheries conservation and management. Fisheries, 44(5): 225–238.
Sass G.G. 2001. An analysis of walleye, Stizostedion vitreum, growth in the Ceded Territory of Wisconsin, 1977–2000. Master's Thesis, University of Wisconsin-Madison.
Sass G.G., Allen M.S. 2014. Foundations of fisheries science. American Fisheries Society, Bethesda, Maryland.
Sass G.G., Kitchell J.F. 2005. Can growth be used as a surrogate measure of walleye (Sander vitreus) abundance change? Canadian Journal of Fisheries and Aquatic Sciences, 62: 2159–2168.
Sass G.G., Shaw S.L. 2018. Walleye population responses to experimental exploitation in a northern Wisconsin lake. Transactions of the American Fisheries Society, 147(5): 869–878.
Sass G.G., Shaw S.L. 2020. Catch-and-release influences on inland recreational fisheries. Reviews in Fisheries Science & Aquaculture, 28: 211–227.
Sass G.G., Hewett S.W., Beard T.D. Jr., Fayram A.H., Kitchell J.F. 2004. The role of density dependence in growth patterns of ceded territory walleye populations of northern Wisconsin: effects of changing management regimes. North American Journal of Fisheries Management, 24(4): 1262–1278.
Sass G.G., Rypel A.L., Stafford J.D. 2017. Inland fisheries habitat management: lessons learned from wildlife ecology and a proposal for change. Fisheries, 42(4): 197–209.
Sass G.G., Shaw S.L., Rooney T.P., Rypel A.L., Raabe J.K., Smith Q.C., et al. 2019. Coarse woody habitat and glacial lake fisheries in the Midwestern United States: knowns, unknowns, and an experiment to advance our knowledge. Lake and Reservoir Management, 35(4): 382–395.
Sass G.G., Feiner Z.S., Shaw S.L. 2021. Empirical evidence for depensation in freshwater fisheries. Fisheries, 46: 266–276.
Sass G.G., Shaw S.L., Gorne J.A., Godard D., Nietlisbach N., Giehtbrock D., et al. 2022a. Female sex ratio bias in extended growth hatchery Walleye fingerlings produced in Wisconsin. North American Journal of Aquaculture, 84: 267–274.
Sass G.G., Shaw S.L., Renik K.M. 2022b. Celebrating 75 years of Wisconsin's Northern Highland Fishery Research Area: the past, present, and future. Fisheries, 47: 55–67.
Sass G.G., Shaw S.L., Sikora L.W., Lorenzoni M., Luehring M. 2022c. Plasticity in abundance and demographic responses of walleye to elevated exploitation in a north temperate lake. North American Journal of Fisheries Management, 42(3): 535–548.
Sass G.G., LaMarche S.T., Feiner Z.S. 2023. Do angler catch and harvest rates differ between open water and ice anglers in Wisconsin? Fisheries Research, 263: 106678.
Schueller A.M., Hansen M.J., Newman S.P. 2008. Modeling the sustainability of walleye populations in northern Wisconsin lakes. North American Journal of Fisheries Management, 28(6): 1916–1927.
Scrogin D., Berrens R.P., Bohara A.K. 2000. Policy changes and the demand for lottery-rationed big game hunting licenses. Journal of Agricultural and Resource Economics, 501–519.
Shaw S.L., Sass G.G., VanDeHey J.A. 2018. Maternal effects better predict walleye recruitment in Escanaba Lake, Wisconsin, 1957–2015: implications for regulations. Canadian Journal of Fisheries and Aquatic Sciences, 75: 2320–2331.
Shaw S.L., Sass G.G., Eslinger L.D. 2019. Effects of angler harvest on adult muskellunge growth and survival in Escanaba Lake, Wisconsin, 1956–2016. North American Journal of Fisheries Management, 39(1): 124–134.
Shaw S.L., Sass G.G., Gorne J. 2023. Identifying ecological factors that influence walleye recruitment in Ceded Territory lakes. Wisconsin Department of Natural Resources, Office of Applied Science, Final Report.
Shultz A., Luehring M., Ray A., Rose J.D., Croll R., Gilbert J., et al. 2022. Case study: applying the resist–accept–direct framework to an Ojibwe Tribe's relationship with the natural world. Fisheries Management and Ecology, 29: 392–408.
Sikora L.W., VanDeHey J.A., Sass G.G., Matzke G., Preul M. 2021. Fish community changes associated with bullhead removals in four northern Wisconsin lakes. North American Journal of Fisheries Management, 41: S71–S81.
Staggs M.D., Moody R.C., Hansen M.J., Hoff M.H. 1990. Spearing and sport angling for walleye in Wisconsin's ceded territory.
Stavins R.N. 2011. The problem of the commons: still unsettled after 100 years. American Economic Review, 101(1): 81–108.
Sullivan M.G. 2002. The illegal harvest of walleye protected by size limits in Alberta. North American Journal of Fisheries Management, 22: 1053–1063.
Tingley R.W. III, Paukert C., Sass G.G., Jacobson P.C., Hansen G.J., Lynch A.J., Shannon P.D. 2019. Adapting to climate change: guidance for the management of inland glacial lake fisheries. Lake and Reservoir Management, 35(4): 435–452.
Tsehaye I., Roth B.M., Sass G.G. 2016. Exploring optimal walleye exploitation rates for northern Wisconsin Ceded Territory lakes using a hierarchical Bayesian age-structured model. Canadian Journal of Fisheries and Aquatic Sciences, 73: 1413–1433.
U.S. Department of the Interior. 1991. Casting light upon the waters: a joint fishery assessment of the Wisconsin Ceded Territory. Bureau of Indian Affairs, Minneapolis, Minnesota.
van Poorten B.T., Camp E.V. 2019. Addressing challenges common to modern recreational fisheries with a buffet-style landscape management approach. Reviews in Fisheries Science & Aquaculture, 27(4): 393–416.
van Poorten B.T., Walters C.J., Ward H.G. 2016. Predicting changes in the catchability coefficient through effort sorting as less skilled fishers exit the fishery during stock declines. Fisheries Research, 183: 379–384.
Vasquez B.R. 2024. Evaluating walleye (Sander vitreus) thermal and optical habitat occupancy in northern Wisconsin lakes using two forms of technology. Master's thesis, University of Wisconsin-Stevens Point.
Ward H.G.M., Askey P.J., Post J.R. 2013. A mechanistic understanding of hyperstability in catch per unit effort and density-dependent catchability in a multistock recreational fishery. Canadian Journal of Fisheries and Aquatic Sciences, 70: 1542–1550.
Walters C., Kitchell J.F. 2001. Cultivation/depensation effects on juvenile survival and recruitment: implications for the theory of fishing. Canadian Journal of Fisheries and Aquatic Sciences, 58(1): 39–50.
Wilkinson G.M., Carpenter S.R., Cole J.J., Pace M.L., Yang C. 2013. Terrestrial support of pelagic consumers: patterns and variability revealed by a multilake study. Freshwater Biology, 58(10): 2037–2049.
Wilson K.L., Foos A., Barker O.E., Farineau A., De Gisi J., Post J.R. 2020. Social–ecological feedbacks drive spatial exploitation in a northern freshwater fishery: a halo of depletion. Journal of Applied Ecology, 57(2): 206–218.
Zebro L.R., Mrnak J.T., Shaw S.L., Chipps S.R., Sass G.G. 2022. Density dependent and environmental influences on juvenile walleye Sander vitreus (Mitchill) survivorship in northern Wisconsin lakes. Fisheries Management and Ecology, 29(6): 897–910.
Information & Authors
Information
Published In

FACETS
Volume 10 • 2025
Pages: 1 - 17
Editor: Eva C Enders
History
Received: 27 March 2024
Accepted: 19 November 2024
Version of record online: 6 February 2025
Copyright
© 2025 The Author. This work is licensed under a Creative Commons Attribution 4.0 International License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Data Availability Statement
Data will be made available upon request.
Key Words
Sections
Subjects
Authors
Author Contributions
Conceptualization: GGS
Funding acquisition: GGS
Writing – original draft: GGS
Writing – review & editing: GGS
Competing Interests
The author declares there are no competing interests.
Funding Information
Wisconsin Department of Natural Resources (WDNR)
Metrics & Citations
Metrics
Other Metrics
Citations
Cite As
Greg G. Sass. 2025. The decline of walleye populations: an ecological tipping point?. FACETS.
10: 1-17.
https://doi.org/10.1139/facets-2024-0064
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
There are no citations for this item