An ensemble machine learning bioavailable strontium isoscape for Eastern Canada
Abstract
Bioavailable strontium isotope ratios (87Sr/86Sr) distribution across the landscape mainly follow the underlying lithology, making 87Sr/86Sr baseline maps (isoscapes) powerful tools for provenance studies. 87Sr/86Sr has already been used in Eastern Canada (EC) to track food and human remains origins, or to reconstruct animal mobility. While bioavailable 87Sr/86Sr isoscapes for EC can be extrapolated from global datasets using random forest modelling (RF), no regionally calibrated isoscape exists. Here, we produce a regionally calibrated bioavailable 87Sr/86Sr isoscape by analysing plants collected at 136 sites across EC, incorporating updated geological variables and applying a novel ensemble machine learning (EML) framework. We generated and compared isoscapes generated by the traditional RF and the EML approaches. Adding local bioavailable 87Sr/86Sr to a global dataset significantly improved the model prediction with a drastic increase of predicted 87Sr/86Sr and increased spatial uncertainty in the northern Canadian craton. EML produced similar 87Sr/86Sr predictions but with tighter spatial uncertainty distribution. Regionally calibrated RF and EML isoscapes significantly outperformed the global bioavailable RF isoscape, confirming the requirement for collecting local data in data-poor regions. This isoscape provides a baseline in EC to monitor and manage the movements and provenance of agricultural products, natural resources, endangered/harmful migratory species, and archaeological human remains and artifacts.
1. Introduction
Strontium (Sr), and more specifically the isotope ratio 87Sr/86Sr, is commonly used as an indicator of geographic origin and mobility in multiple fields of research (Bentley 2006; Baffi and Trincherini 2016; Crowley et al. 2017; Kamenov and Curtis 2017). Mainly driven in the environment by the nature of the underlying lithology through the weathering of the bedrock, the spatial distribution of bioavailable 87Sr/86Sr (i.e., accessible for uptake by living organisms) across the landscape, displays stable scalar patterns that vary little at human timescales (Capo et al. 1998). This stability and the unique mode of spatial variations of bioavailable 87Sr/86Sr have made it a tool of choice for provenance studies. In archaeology, bioavailable 87Sr/86Sr are often used to determine the origin of animal and human remains at archaeological sites and to determine whether the studied individuals were local or not. Such information can thus illuminate personal and group mobility, trading habits, origins, and interactions of past human populations (Alt et al. 2014; Coutu et al. 2016; Shaw et al. 2016; Madgwick et al. 2019a; Britton and Le Corre 2022; Czére et al. 2022; Serna et al. 2023). Bioavailable 87Sr/86Sr data are also useful in palaeoecological case studies to track the mobility of extant and extinct species unearthed at archaeological or palaeontological sites (Britton et al. 2011, 2023; Gigleux et al. 2019; Wooller et al. 2021) and in modern ecology to follow the migratory movements and breeding grounds of birds, mammals, and insects (Font et al. 2007; Crowley et al. 2017, 2021; Miller et al. 2021; Reich et al. 2021). Lastly, bioavailable 87Sr/86Sr datasets have also shown promise in the application of 87Sr/86Sr analysis to certify, authenticate, and trace the origin of foods (Baffi and Trincherini 2016) and forensic materials (Kamenov and Curtis 2017). In parallel to the growing interest in, and applications of, 87Sr/86Sr analysis in provenance studies, the tools and methods to map bioavailable 87Sr/86Sr across the landscape have progressed (Beard and Johnson 2000; Bataille and Bowen 2012; Bataille et al. 2018, 2020, 2021; Funck et al. 2021; Holt et al. 2021). While global bioavailable 87Sr/86Sr predictions now exist and can be used for provenance, several studies have demonstrated that global bioavailable 87Sr/86Sr isoscapes, a map of the spatial variations in isotopic values, show poor predictive power in data-poor regions (Bataille et al. 2020; Wang et al. 2023). Consequently, there is a major incentive to generate more bioavailable 87Sr/86Sr data and to re-calibrate bioavailable strontium isoscapes at smaller scales and in data-poor regions.
The sole natural radiogenic isotope of strontium, 87Sr, is the product of the radioactive decay of 87Rb (rubidium). Due to this decay, the 87Sr abundance in rocks increases through time. Furthermore, ancient rocks with an initially high content of 87Rb, such as granite and rhyolite, are expected to have high 87Sr/86Sr (Bentley 2006; Bataille et al. 2020). Conversely, younger rocks and (or) rocks with low 87Rb content, such as basalt or carbonate, are expected to have a lower 87Sr/86Sr (Bentley 2006; Bataille et al. 2020). Through weathering and leaching, bedrock contributes as the main input to the 87Sr/86Sr available in the soil for plants through pedogenesis (Bentley 2006), alongside lesser inputs, such as sea spray, rainfall, or aeolian deposition (Crowley et al. 2017; Bataille et al. 2020). Bioavailable 87Sr/86Sr is then absorbed by plants or ingested by animals through food and water consumption. In animals, strontium substitutes for calcium and is incorporated into hard tissues such as bones, tooth enamel, otoliths, and shells during growth and mineralisation (Dahl et al. 2001; Willmes et al. 2016; El Meknassi et al. 2018; Guiserix et al. 2022). During this process, 87Sr/86Sr shows very low fractionation (Guiserix et al. 2022), allowing for mass-dependent fractionation correction during analysis. Thus, 87Sr/86Sr in plant and animal tissues reflects bioavailable 87Sr/86Sr, and direct geographic inferences from the 87Sr/86Sr of a given sample can be made with reference to bioavailable 87Sr/86Sr.
In recent years, multiple methods have been used to map variability in the 87Sr/86Sr of environmental samples (e.g., soils, plants) across landscapes and to produce isoscapes at different scales, mapping mean predictions and associated spatial uncertainty (Bataille et al. 2018; Holt et al. 2021). The most straightforward approach is to infer 87Sr/86Sr directly from biological and soil samples. Strontium isotope ratios are 87Sr/86Sr is estimated in samples collected across an area of interest and used to obtain an average 87Sr/86Sr within specific sub-areas of known differences in their geological and lithological characteristics (Evans et al. 2010; Snoeck et al. 2020). Samples of interest (e.g., bones, archaeological remains, food products) are then compared to the previously generated spatial distribution of 87Sr/86Sr to determine whether they are local or not, and (or) to identify their origin (Vinciguerra et al. 2016; Czére et al. 2022; Dargent et al. 2023). Also, relying directly on biological and soil samples, geostatistical approaches, such as kriging with external drift, can be used to interpolate bioavailable 87Sr/86Sr between sampling sites to produce 87Sr/86Sr continuous distribution surfaces, accounting for the discrete distribution of the lithological units in the landscape (Willmes et al. 2018; Britton et al. 2020; Lugli et al. 2022). Kriging also generates spatial uncertainty maps, allowing, in combination with the 87Sr/86Sr continuous distribution surfaces, for probabilistic geographic assignment of the samples of interest (Willmes et al. 2018; Britton et al. 2020; Lugli et al. 2022). A major limitation of direct inference and geostatistical approaches is that they require an extensive sampling effort to obtain well-defined isoscapes (Bataille et al. 2018; Holt et al. 2021) and large-scale isoscapes need to rely on large environmental 87Sr/86Sr databases (Willmes et al. 2018). Moreover, they are limited to the areas delineated by the sampling sites (Holt et al. 2021). As such, these approaches may not be suitable in regions where extensive sampling is not feasible due to cost, accessibility, or scale.
Alternatively, process-based approaches have been proposed to predict the spatial distribution of 87Sr/86Sr at large scales (Bataille and Bowen 2012; Bataille et al. 2020). Bataille et al. (2014) developed a mechanistic model to predict 87Sr/86Sr in bedrock according to bedrock age and lithology. The model relies on the known rate of radioactive decay of the 87Rb into 87Sr and, as a consequence, the increase in 87Sr/86Sr of rocks through time. In such models, equations typically integrate bedrock age and the 87Rb/86Sr ratio of actual and parent rock material (Bataille and Bowen 2012; Bataille et al. 2014). While these models can provide a strong baseline 87Sr/86Sr bedrock isoscape, they may fail to reflect the bioavailable 87Sr/86Sr spatial distribution, particularly for old and heterogeneous bedrock or in areas where other sources of bioavailable Sr (e.g., precipitation, aerosol deposits, fertiliser) are more influential (Bataille et al. 2020; Holt et al. 2021). Building on their mechanistic bedrock model, Bataille et al. (2018) proposed a model based on a random forest regression (RF, Breiman 2001), machine learning algorithm that integrates the process-based 87Sr/86Sr bedrock isoscape and geospatial variables representing the potential auxiliary sources of Sr to ecosystems together into a regression. The model is trained on a dataset of bioavailable 87Sr/86Sr from different analytes (i.e., plants, local animals, exchangeable soil Sr, water), compiled by Bataiille et al. (2018, 2020), and predicts bioavailable 87Sr/86Sr, alongside spatial uncertainty maps (Bataille et al. 2018, 2020). This approach allows the predictions of bioavailable 87Sr/86Sr at very large scales and in areas with low or no environmental sample coverage. However, Bataille et al. (2020) also cautioned that this RF approach shows a limited power of extrapolation when predicting bioavailable 87Sr/86Sr in regions without training data (Bataille et al. 2020; Wang et al. 2023). This caveat is especially true when predicting bioavailable 87Sr/86Sr for which the geology, climate, or environments that are not well represented in the training dataset. For example, Wang et al. (2023) showed that the global predictions of bioavailable 87Sr/86Sr isoscape had very low accuracy in Angola, likely because the compiled training database has almost no samples for the old radiogenic cratonic regions and tropical areas typical of this region.
While RF generally outperforms other modelling methods to map bioavailable 87Sr/86Sr (Cutler et al. 2007; Bataille et al. 2018), it only relies on a single algorithm to extrapolate predictions. Recent developments in numerical methods for mapping soil properties have emphasised the power of combining algorithms (e.g., ensemble learning), including RF, to improve the unbiasedness, extrapolation potential, and estimates of uncertainty of soil property predictions (Wadoux et al. 2020; Hengl et al. 2021). Another downside of the current RF framework relative to existing interpolation approaches is that it does not account for spatial autocorrelation in the calibration dataset (i.e., the spatial predictions and uncertainty estimates solely depend on geospatial predictors and are not influenced by the location of existing observations). Several approaches have been proposed to solve this issue, including adding geographical features such as buffer or oblique distances (Møller et al. 2020) and combining multi-scale RF models (Georganos et al. 2021).
Eastern Canada (EC) is an excellent location to test novel mapping approaches for bioavailable 87Sr/86Sr. The geology, climate, and environment of this region are unique and not well represented by existing bioavailable 87Sr/86Sr isoscapes (Bataille et al. 2020). Yet, provenance studies using 87Sr/86Sr analysis are growing (e.g., Stevenson et al. 2015; Guibourdenche et al. 2020; Pfeiffer et al. 2020; Fauberteau et al. 2021; Dargent et al. 2023). The geology of EC is dominated in the north by the large Canadian craton composed of a diversity of plutonic and metamorphic geological provinces and units dating from the Archaean and Proterozoic periods (Shilts et al. 1987). Due to its old age and a range of lithology with distinct Rb content, very broad 87Sr/86Sr variations are expected in the bedrock of this region (and thus in that of its soils and plants). Conversely, south of the St. Lawrence River, Palaeozoic marine sedimentary formations, igneous and metamorphic units of the Appalachian mountains, Pleistocene glacial sediments, and Champlain Sea deposits (Thériault and Beauséjour 2012) dominate the landscape and contribute to probably lower but variable bioavailable 87Sr/86Sr in that area. Although incorporating a few data from the European craton, the database of globally compiled bioavailable 87Sr/86Sr mainly consists of samples with bioavailable 87Sr/86Sr below 0.720 and lacks samples from areas of higher bioavailable 87Sr/86Sr such as those originating from old, highly radiogenic regions of the Canadian craton (Bataille et al. 2020). Moreover, the Global Lithological Map (GLiM, Hartmann and Moosdorf 2012) used to build the mechanistic bedrock 87Sr/86Sr model (Bataille et al. 2014), which is the main predictor of bioavailable 87Sr/86Sr, relies on outdated and low-resolution geological data. In addition to its unique geology, EC also shows a unique climate with temperate forests characterised by cold, snowy winters and warm humid summers in southern EC, boreal forests with colder sub-arctic climates in central EC and tundra-like peatland environments in the northern EC. Incorporating bioavailable data from this geologically and environmentally unique region is thus essential to further improve regional and global 87Sr/86Sr isoscapes.
Besides its direct interest as an undersampled testing site to improve the modelling and coverage of bioavailable 87Sr/86Sr, EC is also known for its local agricultural production, thriving forest industry, biodiversity, and the historical presence of Inuit and First Nation communities, which could all benefit from the development of provenance tools. Provenancing using 87Sr/86Sr has shown potential in food sciences and food forensics in the region, for example, to certify the origins of locally produced wine (Vinciguerra et al. 2016; Guibourdenche et al. 2020), cheese (Stevenson et al. 2015) and maple syrup (Nguyen et al. 2023). Similarly, bioavailable 87Sr/86Sr have been used to trace the origin of human remains from their tissues to investigate cold cases (Fauberteau et al. 2021), to trace the origin of historical immigrants in the Montreal region (Vautour et al. 2015), and to reconstruct the history of movements of First Nations people (Pfeiffer et al. 2020). In forestry, the recent application of 87Sr/86Sr in Canada showed its potential to trace the dispersal and population connectivity of forest pests destroying the boreal forest, such as the eastern spruce budworm moth (Choristoneura fumiferana, Dargent et al. 2023). Finally, the large and remote landscape of EC is increasingly affected by human disturbances and warming that are rapidly changing the suitability of breeding grounds, migration routes and habitats of the many mammals, birds and insect species that inhabit this region (Foster et al. 2022). These changes in mobility and habitat suitability are one of the major drivers of the rapid population decline of many of these boreal species (Vors and Boyce 2009; Gauthier et al. 2013). There is therefore an urgent need to develop tools that can allow the generation of modern and historic datasets, such as bioavailable 86Sr/87Sr, to better understand past and present changes in the mobility, range, and seasonal breeding sites of this priceless biodiversity resource.
In this paper, we aimed to develop an accurate, unbiased, and regionally calibrated isoscape for EC by adding new plant samples collected in the region to the global database from Bataille et al. (2020), and by updating the bedrock model and the geological variables for EC with a more recent and accurate lithological map (Thériault and Beauséjour 2012). We also tested a new mapping approach by comparing the traditional RF approach from Bataille et al. (2020) to a spatial interpolation ensemble machine learning (EML) approach (Hengl et al. 2021) to explore how ensemble learning can improve model predictions and spatial uncertainty estimates. We conclude by discussing the potential of this new isoscape for provenance studies in the region.
2. Methods
2.1. Geological context
EC is composed of several geological provinces (Fig. 1a), large-scale regions that share similar geological characteristics. North of the St. Lawrence River, the Canadian craton, dominated by metamorphic and plutonic rocks, is divided into three main geological provinces. The Superior Province (the core of Northern Québec region and covering two-thirds of Ontario) and the Churchill Province (lying North and East of the Superior Province and covering Québec and Labrador) were formed during the Mesoarchaean and Mesoproterozoic (Thériault and Beauséjour 2012). The Grenville Province, south of these two other provinces, was formed during the Archaean in the parautochton region (contiguous to the Superior Province) and also during the Paleo- and Meso-Proterozoic in the allochton region. The Canadian craton also includes smaller geological provinces, with parts of the Southern Province in Ontario (Paleo-Mesoproterozoic) and the Nain Province along the eastern coast of Labrador (Meso-Neoarchaean). Besides the Canadian craton, the Hudson Bay lowlands to the North of Ontario consist mainly of Palaeozoic–Mesozoic carbonate sediments. South of the St. Lawrence River, the Appalachian orogen was formed during the Palaeozoic and is dominated by siliciclastic sediments in southern Québec and New Brunswick and by metamorphic rocks in Newfoundland. Finally, the St. Lawrence Platform, located between the Grenville province and the Appalachian orogen consists of Palaeozoic carbonate sedimentary rocks. The landscape of EC has also been shaped by past glaciations and is marked by the presence of numerous tills, glacial lacustrine sediments, and eskers. The topography of the EC provinces notably reflects the past presence of the Laurentide Ice Sheet (Sugden 1978).
Fig. 1.
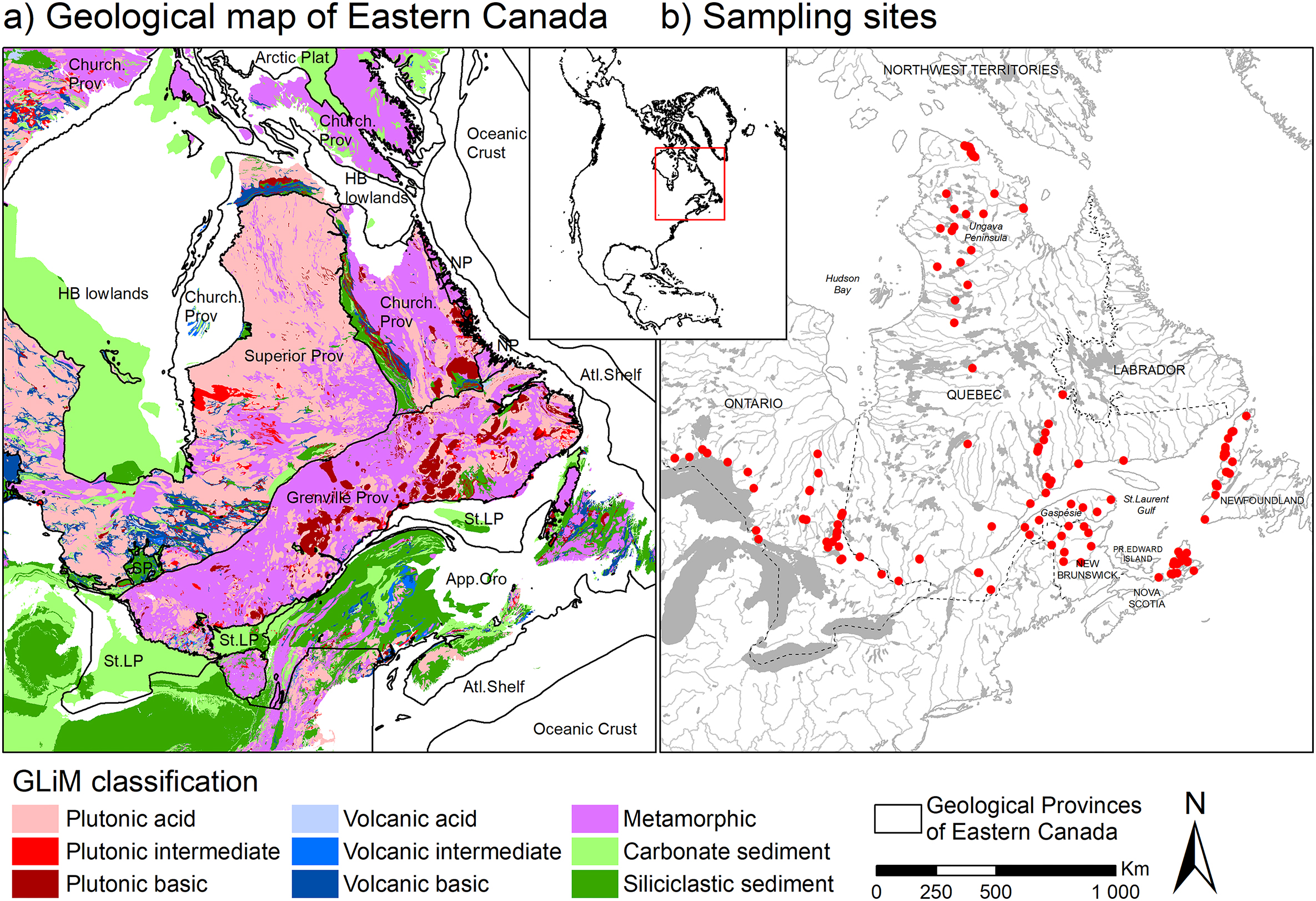
2.2. Sample collection and laboratory protocols
We completed the strontium database already compiled by Bataille et al. (2020) and Funck et al. (2021) with two independent sampling campaigns conducted across EC between 2018 and 2022 (Fig. 1b). Opportunistic and planned vegetation sampling were conducted by the Caribou Ungava research program (Université Laval, Québec) in northern Québec, alongside their ongoing field activities. Plants (moss, lichen and grass) were collected at 28 sites across taiga and tundra habitats. Plant samples were prepared and analysed at the Memorial Applied Archaeological Science laboratory, Earth Resources Research and Analysis Facility, and Microanalysis Facility in the CREAIT Network, Memorial University of Newfoundland, Canada. Plant samples were processed using a modified version of the approach described in Britton et al. (2020), and strontium was then extracted from the samples using a modification of the method from Deniel and Pin (2001) as detailed in (Copeland et al. 2008). Plant samples were kept in unsealed polyethylene bags following collection and cold-stored until analysis. At Memorial, these were then washed in ultra-pure MilliQ water to remove dust or any other adhering materials, frozen, and freeze-dried. Dried plant samples (≈200 mg) were then ashed in a muffle furnace overnight at 500 °C. Sub-samples of ashed plant material (10–20 mg) were then dissolved in closed 7 mL Savillex perfluoroalkoxy (PFA) vials (Minnetonka, MN, USA) with 2 mL 8 mol/L HNO3 (single distilled Reagent grade, Sigma Aldrich) at 120 °C for 48 h. After cooling to room temperature, 1 mL of high purity 30% H2O2 was added and the vials heated to 90 °C for 24 h. The clear solutions were then evaporated at 90 °C and 1 mL of 8 mol/L HNO3 was added to redissolve residual material. The 1 mL 8 mol/L HNO3 solution was then transferred into pre-conditioned 1 mL microcolumns (after Charlier et al. 2006) containing ≈ 250 µL Sr-spec Resin™ (100–150 µm particle size, EiChrom, Lisle, IL, USA), washed three times with 1 mL of 8 mol/L HNO3, and strontium was eluted from the resin in 2 mL MilliQ water then acidified with the addition of 75 µL of 8 mol/L HNO3. Analysis of 87Sr/86Sr in the purified Sr solutions was conducted using a Neptune Multi-Collector Inductively Coupled Plasma Mass Spectrometer (MC-ICP-MS, Thermo Fisher Scientific, Bremen, Germany) fitted with a 50 µL/min PFA nebulizer using instrument settings and analytical methods typical for solution Sr isotope analyses (Copeland et al. 2008; Madgwick et al. 2019b). A 200 ppb solution of Sr isotope standard SRM 987 (strontium carbonate dissolved in 0.3 mol/L HNO3, National Institute of Standards and Technology, Gaithersburg, MD, USA) was analysed between every five samples for quality control purposes. Using the average 87Sr/86Sr for the SRM 987 collected in the analytical session, all data were externally corrected to the SRM 987 87Sr/86Sr = 0.710240 (Johnson and Fridrich 1990), and the facility's average for SRM 987 in a typical plant sample analytical session (e.g., 15 November 2019) was 0.710294 ± 0.000010 (1SD, n = 11). We assumed no dissimilarity in 87Sr/86Sr between moss and lichen samples compared with vascular plant samples.
Planned conifer needle sampling was conducted across EC and the Maritime provinces as part of the Healthy Forest Partnership Early Intervention Strategy against spruce budworm (e.g., Johns et al. 2019; MacLean 2019). Balsam fir (Abies balsamea (Mill.)), and spruce needles (Picea sp.) were collected at 107 sites across the boreal forest (Fig. 1). At each site of collection, three independent trees were sampled, except for two sites in Quebec and 14 sites in Cape Breton, Nova Scotia, where only samples from a single tree could be obtained, and stored in paper bags until analysis. In the laboratory at the University of Ottawa, 0.02 g of needles per tree were sub-sampled and aggregated by site. Conifer needles were then cleaned through sonication in distilled de-ionized water for 10 min to remove surface dust, after which the needles were dried in an oven at 50 °C for 48 h. Approximately 0.06 g of needles, per site, were then digested in 1 mL concentrated nitric acid (16 mol/L HNO3, TraceMetal™ grade; Fisher Chemical, Canada) in PFA tubes at 250 °C for 15 min using microwave digestion (Organic High setting—Anton Parr Multiwave 7000, Austria). Following digestion, the liquid was clear, suggesting complete digestion. We placed the digested liquid into 7 mL Savillex PTFE. An aliquot of 50 µL of the solution from each Savillex vial was pipetted to Labcon MetalFree™ centrifuge tubes and diluted with 2 mL of 2% v/v HNO3. Sr concentration analysis was performed by ICP-MS (Agilent 8800 triple quadrupole mass spectrometer) at the Department of Earth and Environmental Sciences, University of Ottawa. Calibration standards were prepared using single element certified standards purchased from SCP Science (Montreal, Canada). The remaining ∼1 mL aliquot of the sample in the 7 mL Savillex PTFE vial was dried down and re-dissolved in 1 mL 6 mol/L HNO3. The separation of Sr was processed in 100 µL microcolumns loaded with Sr-spec Resin™ (100–150 µm; Eichrom Technologies, LLC). The matrix was rinsed out using 6 mol/L HNO3. The Sr was collected with 0.05 mol/L HNO3. After separation, the eluates were dried and re-dissolved in 200 µL 2% v/v HNO3 for 87Sr/86Sr analysis. The 87Sr/86Sr analysis was performed at the Pacific Centre for Isotopic and Geochemical Research using a Nu-Plasma II high-resolution MC-ICP-MS (Nu Instruments) coupled to a desolvating nebulizer (Aridus II™, CETAC Technologies). The reproducibility of the 87Sr/86Sr measurement for 5 ppb NIST SRM 987 is 0.71025 ± 0.00009 (1 SD, n = 138).
By incorporating the newly analysed samples from both collection initiative described above, the database counted 9161 samples from 4677 sites. 87Sr/86Sr analysed in this study are provided in Supplementary material Table S1.
2.3. Mechanistic bedrock model
We updated the mechanistic bedrock model used in (Bataille et al. 2020) for the province of Québec, using SIGEOM (Système d'Information Géominière, Ministère des Ressources naturelles et des Forêts (MRNF), Québec) fine scale geological data (Thériault and Beauséjour 2012). The mechanistic bedrock model was developed in (Bataille et al. 2014). To estimate modern 87Sr/86Sr of rocks, the model assumes that all rocks of a given lithology come from a common parent material. From its parent material, the 87Sr/86Sr of the modern rock changed over time following a three-stage history with (1) the initial 87Sr/86Sr in undifferentiated mantle, (2) the enrichment in 87Sr through 87Rb decay of the parent crustal rock reservoir from its differentiation to the formation of the modern rock, and (3) the enrichment in 87Sr of the modern rock since its formation. At each stage, the lithology inherits the 87Sr/86Sr of its parent material but with a different chemical composition in Rb and Sr, leading to a different enrichment rate according to the stage. The model is divided into three sub-models for igneous and metamorphic rocks, siliciclastic sediments, and carbonate sediments. It accounts for the rock type and age of each geological unit as well as geochemical data (Rb/Sr) of these units to predicted the 87Sr/86Sr of the modern bedrock. Details and equations of the model are provided in Supplementary material SM2.
The SIGEOM database provides shapefiles with rocks type and age description of the lithological units (Thériault and Beauséjour 2012; Ministère des Ressources naturelles et des Forêts du Québec (MRNF) 2021a) along with geochemical data of rock samples (MRNF 2021b) from all over the province of Québec. Following Bataille et al. (2014), the age descriptors were converted into numerical values, based on the geological timescale, to obtain the minimum and maximum ages of each geological unit. Lithological units were regrouped into nine categories according to their mineral composition following the GLiM classification (Hartmann and Moosdorf 2012): (1) plutonic acid/felsic (pa), (2) plutonic intermediate (pi), (3) plutonic basic/mafic rocks (pb), (4) volcanic acid/felsic (va), (5) volcanic intermediate (vi), (6) volcanic basic/mafic rocks (vb), (7) metamorphic rocks (mt), (8) siliciclastic sediments (ss), and (9) carbonate sediments (ca). We extracted the Rb/Sr data from the geochemical dataset sampled across Québec and estimated spatial variation of modern 87Rb/86Sr within each of the GLiM units using ordinary kriging. We then applied the mechanistic bedrock model from Bataille et al. (2014) to generate 87Sr/86Sr bedrock isoscapes corresponding to the median, 1st quartile, and 3rd quartile of the 87Sr/86Sr prediction. Finally, we updated the 87Sr/86Sr bedrock isoscapes rasters and the geological age rasters from Bataille et al. (2020) with the new data generated for Québec. Kriging was done in ArcGIS (v10.3), and the mechanistic bedrock model was run in R (R Core Team 2023). The R script, adapted from Bataille et al. (2014), is available as Supplementary material SM3.
2.4. Auxiliary variables
In addition to the 87Sr/86Sr bedrock isoscapes (median, 1st quartile, 3rd quartile, Fig. S1) and minimal/maximal age of the bedrock, we used covariates that were likely to influence the bioavailable 87Sr/86Sr in the environment to build the process-based model (Bataille et al. 2020). These variables included terrane age, topography, soil properties, climate, and salt and dust deposits. Although tested by Bataille et al. (2020), we did not include fertiliser variables while fertilisers are expected to be the main anthropogenic influence on bioavailable strontium in soil 87Sr/86Sr (Maurer et al. 2012). We discarded this variable as EC is mainly composed of forest, taiga, and tundra habitats with limited agricultural surfaces in the south (Latifovic et al. 2004), and because fertiliser variables were not retained in the global isoscape model of Bataille et al. (2020). The list of the covariates is provided in Table 1. We extracted geologic, environmental, and climate variables for each sampling site. Due to the multiple sources, covariates vary in terms of accuracy, coverage, and resolution, and all variables were not available at each site. In these cases (161/4677 sites), the nearest covariate value in the vicinity of the site was used.
Table 1.
| Variable | Description | Resolution | Reference |
|---|---|---|---|
| r.m1 | Median bedrock model | 1 km | Bataille et al. (2020); this study |
| r.srsrq1 | 1st quartile bedrock model | 1 km | Bataille et al. (2020); this study |
| r.srsrq3 | 3rd quartile bedrock model | 1 km | Bataille et al. (2020); this study |
| r.meanage_geol | Mean GLiM age (Myr) | 1 km | Hartmann and Moosdorf (2012); this study |
| r.minage_geol | Minimal GLiM age (Myr) | 1 km | Hartmann and Moosdorf (2012); this study |
| r.maxage_geol | Maximal GLiM age (Myr) | 1 km | Hartmann and Moosdorf (2012); this study |
| r.age | Terrane age (Myrs) | 1 km | Mooney et al. (1998) |
| r.mat | Mean annual temperature (°C) | 30 arc sec | Hijmans et al. (2005) |
| r.map | Mean annual precipitation (mm.year−1) | 30 arc sec | Hijmans et al. (2005) |
| r.pet | Global potential evapo-transpiration | 30 arc sec | Zomer et al. (2008) |
| r.ai | Global aridity index | 30 arc sec | Zomer et al. (2008) |
| r.salt | Simulation of sea salt deposition (g.m−2.year−1) | 1° × 1° | Vet et al. (2014) |
| r.dust | Dust deposition (g.m−2.year−1) | 1° × 1° | Mahowald et al. (2006) |
| r.elevation | Shuttle radar topography mission (m) | 90 m | Jarvis et al. (2008) |
| r.bouguer | Bouguer anomaly | 2 min | Balmino et al. (2012) |
| r.GUM | Global unconsolidated sediment map | 1 km | Börker et al. (2018) |
| r.cec | Cation exchange capacity (mmol(c)/kg) | 250 m | Poggio et al. (2021) |
| r.ph | Soil pH (H2O, x10) | 250 m | Poggio et al. (2021) |
| r.phkcl | Soil pH (KCl, x10) | 250 m | Poggio et al. (2021) |
| r.clay | Clay (g/kg) | 250 m | Poggio et al. (2021) |
| r.ocs | Organic carbon stocks (t/ha) | 250 m | Poggio et al. (2021) |
| r.bulk | Bulk density (cg.cm−3) | 250 m | Poggio et al. (2021) |
Note: GLiM: Global Lithological Map.
2.5. Random forest
RF is a machine learning algorithm based on regression trees (Breiman 2001). Multiple regression trees are grown by bagging: for each tree, the dataset is divided randomly by bootstrap into a training set and a validation set, or “out-of-bag”. The training set is used to grow the tree on a random subset of the predictors, and the “out-of-bag” set is used for internal cross-validation. Predictions are then obtained by aggregating the outcome of each tree. RF does not require assumptions on data distribution or variance homogeneity and is resilient to overfitting (Breiman 2001). Analyses were conducted in R (v4.3), and the script, adapted from Bataille et al. (2020), is available as Supplementary material SM4.
Before running the RF analysis, we proceeded to a variable selection step. First, we removed strongly correlated variables (R > 0.9). Random forests are marginally affected by collinearity between covariates (Cutler et al. 2007), but a very high correlation between two variables may artificially increase their influence on the model (Strobl et al. 2008). We then conducted a variable selection using the VSURF R package (Genuer et al. 2015), a three-step algorithm that detects and removes irrelevant and redundant variables in RF models. We used the covariates retained to run the final model, setting the number of trees to be grown to 3000 (Breiman 2001). The performance of the model was evaluated using the root-mean-square error (RMSE) and a 10-fold cross-validation repeated five times. Variable importance, an estimate of how a given variable improve the performance of the model, was determined by node impurity, which reflects the efficiency of the trees to split the data in two homogeneous group.
Geographic assignment on continuous-probability surface models requires assessing the spatial uncertainty of the isoscapes (Wunder 2012). We used quantile RF to calculate a 68.27% prediction interval (Funck et al. 2021) and estimated the standard deviation (σ) from its lower and upper limits:
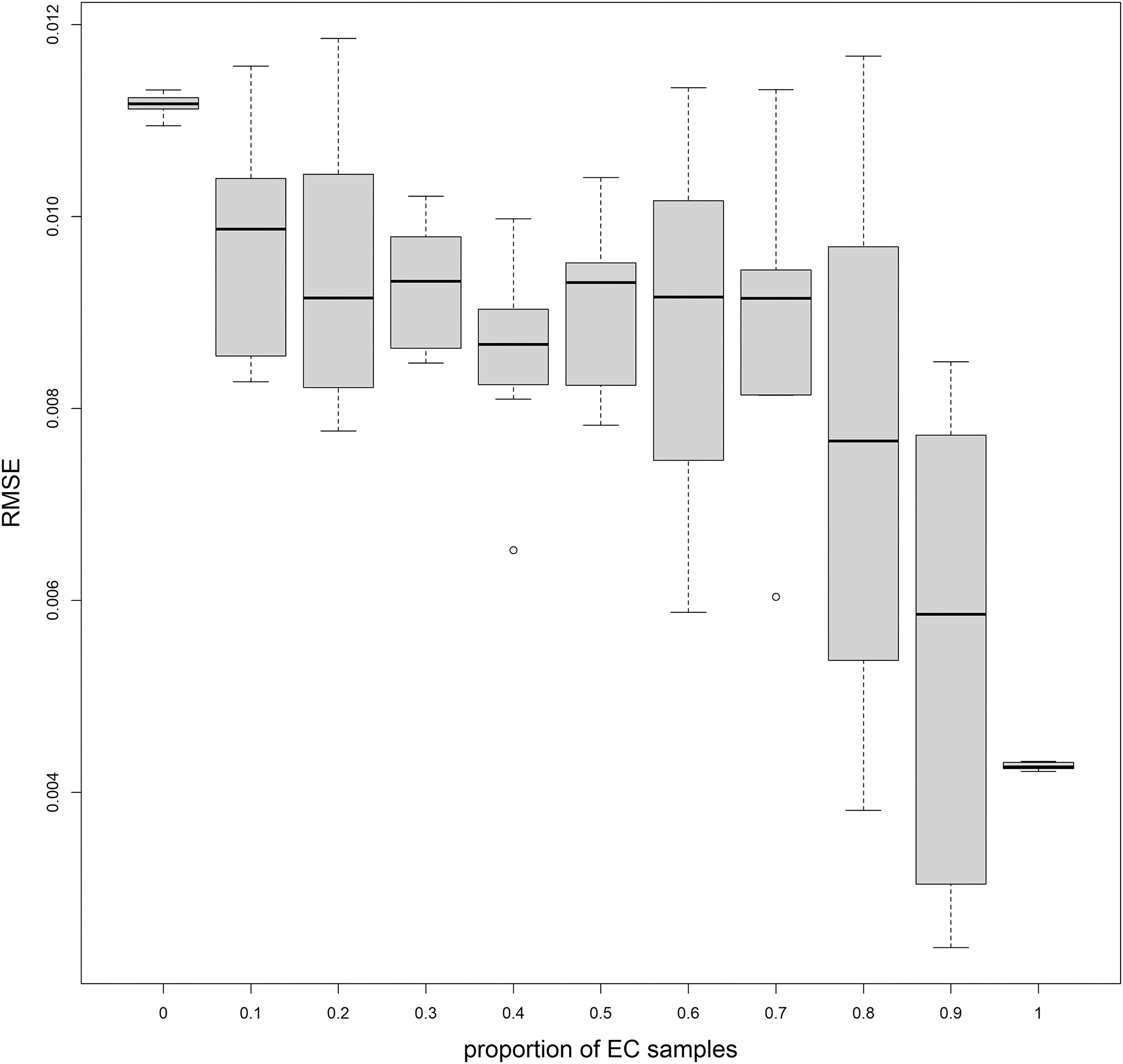
The RF analyses were conducted with the samples from EC (RFalldata) and without them (RFnoEC). We also tested how adding samples to the global dataset improved local predictions. We ran the RFalldata model using from 0% to 100% of the EC dataset, with a 10% increment, selected randomly, and we calculated RMSE for the remaining EC samples, except when 100% of the EC was used. In this case RMSE was calculated on the complete EC dataset. The analysis was repeated 10 times for each proportion.
2.6. Spatial interpolation ensemble machine learning
EML is a set of methods that combine the predictions and outcomes of multiple individual models (i.e., learners) to reduce the error and increase the accuracy of the predictions (Zhang and Ma 2012). Among the different EML methods, the stacking approach aggregates the predictions through the use of a meta-learner. Several learners, based on different algorithms, are trained independently on the same dataset. The predictions of these models are then used to train the meta-learner, thus generating the EML estimates (Zhang and Ma 2012).
We used the landmap R package (Hengl 2023) to apply the stacking approach to our dataset and produce an isoscape on the same extent as the RF model. The landmap package accounts for spatial autocorrelation by using oblique geographic coordinates as covariates (Møller et al. 2020). The modelling framework further accounts for spatial dependencies by applying a spatial cross-validation approach to account for geographic sampling biases. Additionally, EML combines a series of machine learning algorithms, limiting the biases of using a single algorithm (i.e., RF) while improving the performance and robustness of the model.
When applying the spatial interpolation EML, we used the same training set and predictors selected in the RFalldata model. The EML was trained using default learners set of landmap: the function fit five base learners (RF, gradient boosting, support vector machines, neural networks, Lasso and elastic-net regularised generalised linear models), and then used a linear model for the meta-learner. The performance of the EML is assessed through five-fold cross-validation. Quantile forest regression is used in landmap to compute spatial uncertainty similar to what is done with RF. We changed the default value of the number of trees set for the computation of the quantile forest regression from 85 to 1000 directly in the source code of the function. From the model, we generated a mean prediction map, which represents the best unbiased estimate of bioavailable 87Sr/86Sr ratios and the prediction errors map. The R script is available as Supplementary material SM4.
3. Results
3.1. 87Sr/86Sr in sample sites
87Sr/86Sr from the new sample sites from EC covered a broad range of values (0.7087–0.7768), with distinct values according to the main geological provinces of EC. In the old Superior and Churchill provinces, mean 87Sr/86Sr at sampling sites were high and variable (0.7297 ± 0.0157 SD, n = 39, range: 0.7109–0.7768) with the highest values observed in the Ungava Peninsula. Sites from the younger Appalachian orogeny and St. Lawrence platform showed a lower and less variable 87Sr/86Sr (0.7133 ± 0.0033, n = 58, range: 0.7087–0.7214). Finally, sites from the Grenville and Southern provinces showed an intermediate average 87Sr/86Sr and variability (0.718 ± 0.081, n = 38, 0.7097–0.7430).
3.2. Variable selection
Variables selected for the final RF were similar between RFnoEC and RFalldata with the original dataset and when adding the new EC sample sites. Both models included the 1st quartile of the bedrock's model prediction (r.srsrq1), geological variables (r.age, r.maxage), aerosol deposition (r.dust, r.salt), and climate variables (r.mat, r.pet, r.map) but none of the soil properties variables. The two RF only differed by the inclusion of the minimum GLiM age (r.minage) for RFnoEC. For RFalldata, geological and climate variables appeared to be the main predictors of bioavailable 87Sr/86Sr, followed by the bedrock's model prediction (Fig. 2a). However, partial dependence plots (Fig. 2b), depicting the relationship between predictor variables and the prediction of the model, showed greater variation in predicted 87Sr/86Sr for bedrock model predictions and geological variables, as well as for potential evapo-transpiration (r.pet) and temperatures (r.mat). The predicted 87Sr/86Sr increased with the age of the bedrock (r.maxage_geol, r.age) and its 87Sr/86Sr modelled value (r.srsrq1). Predicted 87Sr/86Sr also increased with increasing potential evapo-transpiration (r.pet), temperature (r.mat) and, to a lower extent, precipitation (r.map). Finally, the relationship with salt (r.salt) and dust deposits (r.dust) was less consistent, with no clear trend. While the order of variables differed for RFnoEC, the partial dependence plots were almost identical (Fig. S2).
Fig. 2.
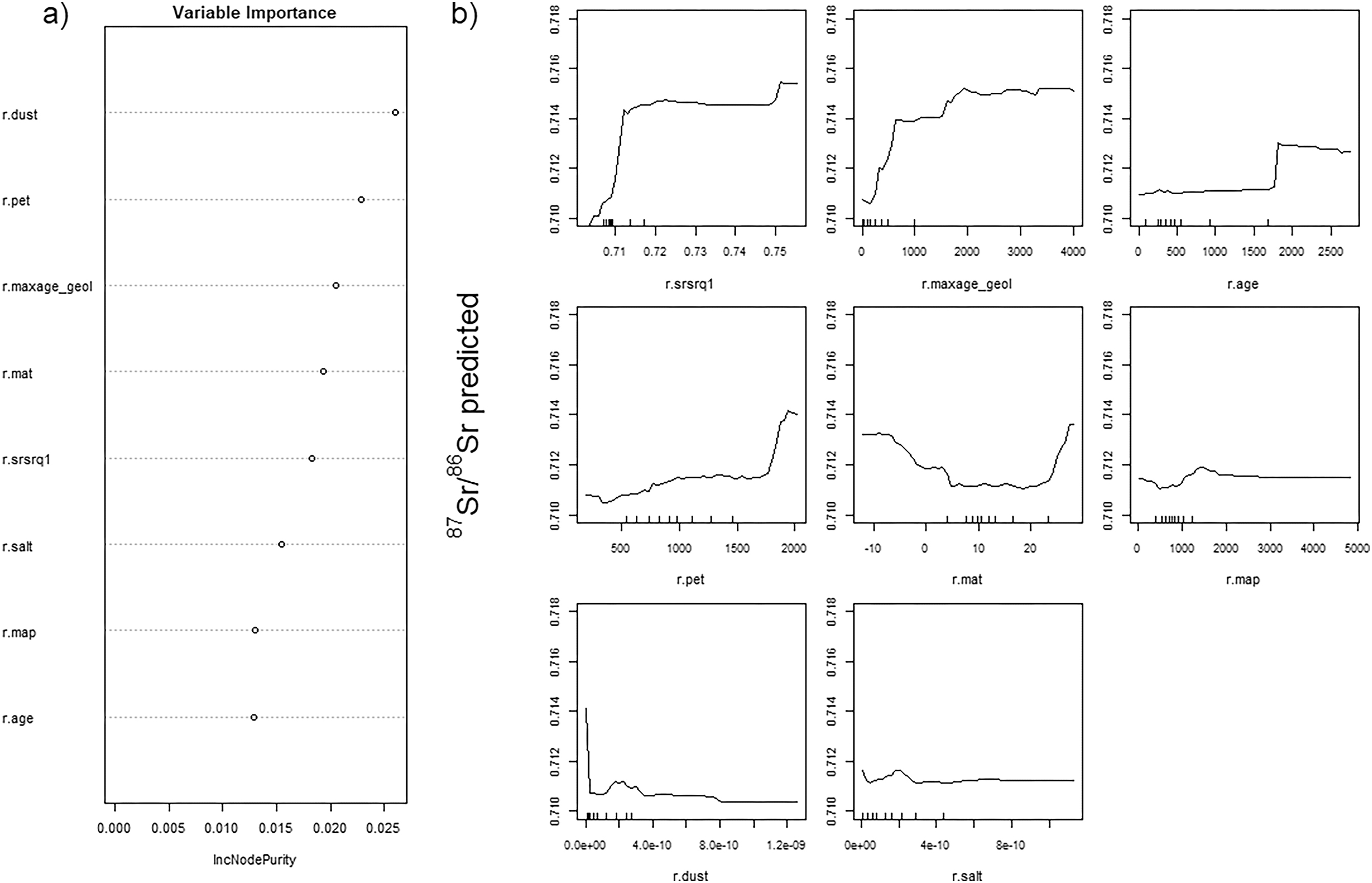
3.3. Performance of the RF and comparison of the isoscapes
Performances of RFnoEC and RFalldata were very similar, and n-fold cross-validation showed a good agreement between predictions and observations (Fig. 3a). RFnoEC explained 61.7% of the variance with an RMSE of 0.0033. Adding EC samples slightly improved the variance explained (63.0%) but increased the RMSE (0.0035). However, looking at local predictions, the RMSE for EC samples only, decreased from 0.0112 (±0.00011) for RFnoEC to 0.0043 (±0.00004) for RFalldata. The bioavailable 87Sr/86Sr isoscapes predicted from the two RF showed also regional differences (Fig. 4). Adding EC samples increased the predicted values of bioavailable 87Sr/86Sr for the old Superior and Churchill geological provinces of Québec and Labrador, and to a lesser extent, in the parautochton part of the Grenville province. For RFnoEC, northern Québec appeared relatively homogeneous with 87Sr/86Sr ranging from 0.715 to 0.726 (Fig. 4a). For RFalldata, predicted 87Sr/86Sr in northern Québec rose above 0.725 (Fig. 4c) with a clear distinction between the southern part (0.725–0.735) and the northern part of the Ungava peninsula (>0.735). However, relative differences in 87Sr/86Sr between geological units remained visible with, for example, lower 87Sr/86Sr in the volcanic Puvirnituq Mountains in the extreme north of Québec (Figs. 1a and 4). Conversely, 87Sr/86Sr ratios of the southern, younger, geological regions (St. Lawrence platform, Appalachian orogen) remained similar for RFnoEC and RFalldata (Figs. 4a and 4c). Adding EC samples also changed the spatial distribution of the error associated with the prediction (Figs. 4b and 4d). In the southern geological provinces, the spatial error slightly decreased but in the northern geological regions, particularly in the Ungava peninsula where the predicted 87Sr/86Sr is the highest, the uncertainty drastically increased reaching in some areas 0.015–0.030 (SD). Finally, bioavailable 87Sr/86Sr ratios of the EC sample sites showed a better correlation with the predicted bioavailable 87Sr/86Sr from RFalldata (R2 = 0.89, Fig. 3b) than with the predicted bioavailable 87Sr/86Sr from RFnoEC (R2 = 0.34). Adding an increasing proportion of the EC dataset to the original dataset improved local prediction (decrease in RMSE) but showed a large variability (Fig. 5).
Fig. 3.
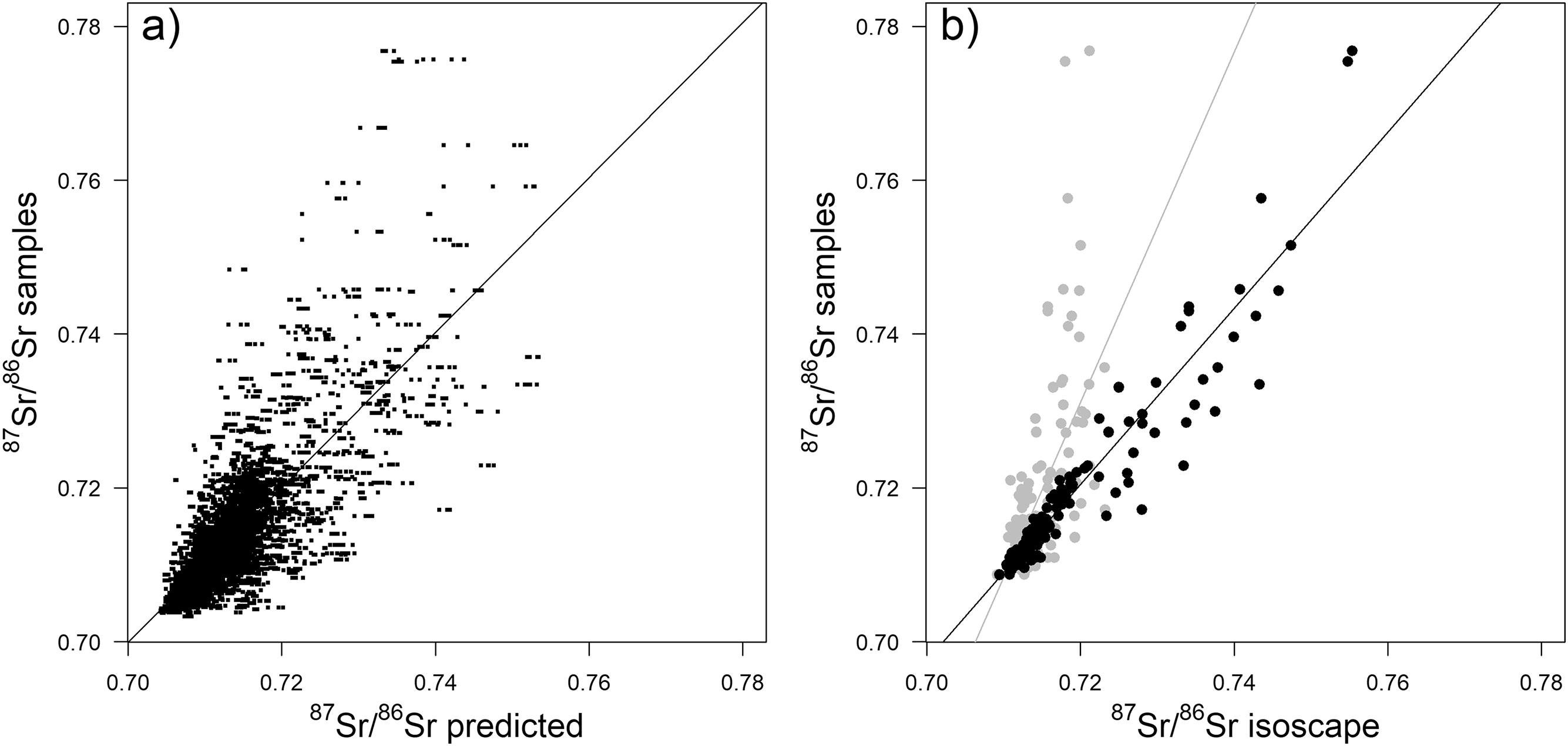
Fig. 4.
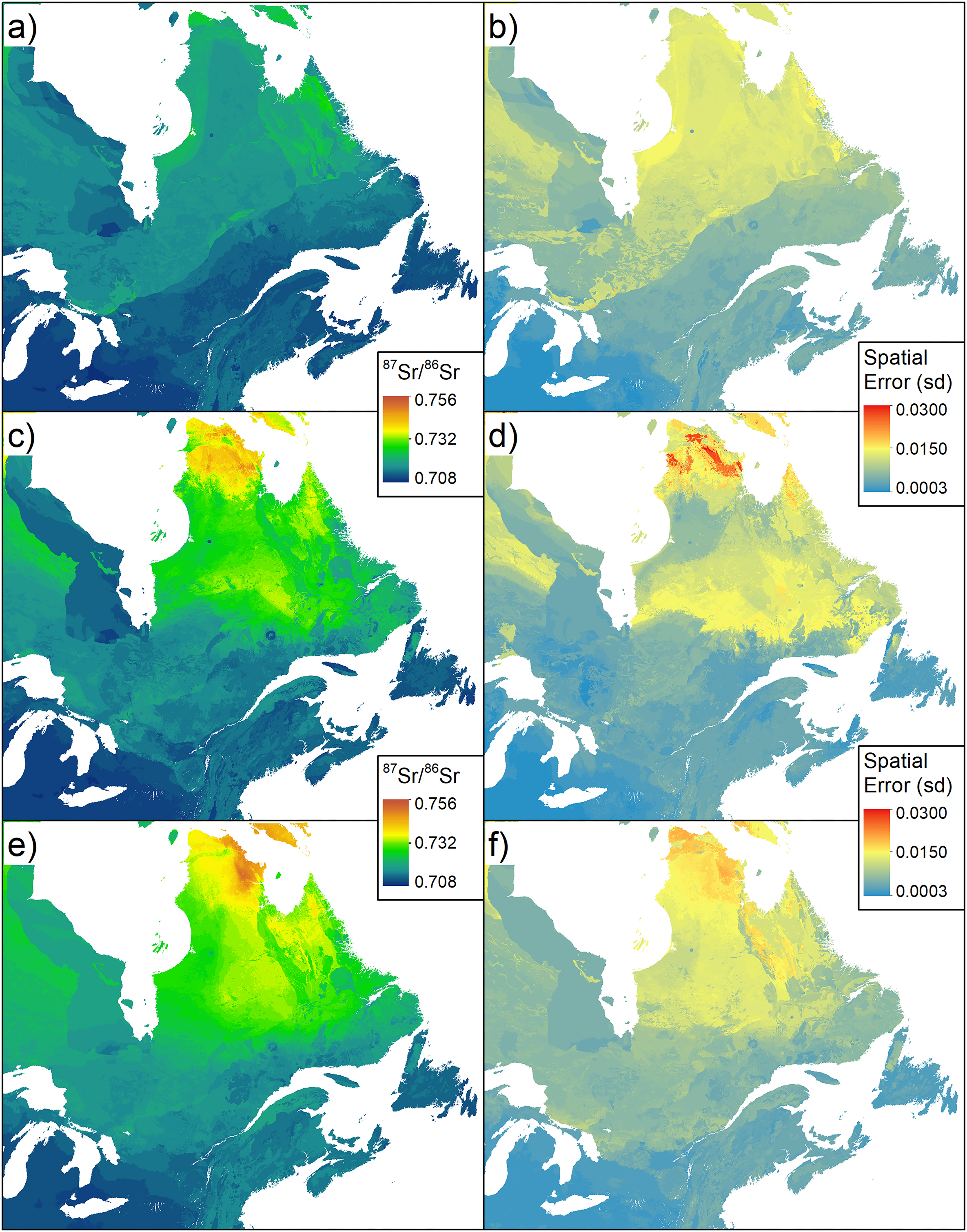
Fig. 5.
3.4. Ensemble machine learning
The variance and RMSE explained by the EML model (EMLalldata) were similar to RFalldata and did not show a significant improvement in accuracy (R2 = 0.66, RMSE = 0.0035). Absolute t values of the learners in the meta-model indicate that RF was the strongest learner (Table 2). Predictions of bioavailable 87Sr/86Sr across the landscape were similar between RFalldata and EMLalldata but with higher bioavailable 87Sr/86Sr in the northeast (Figs. 4c and 4e). However, major differences appeared in spatial uncertainty patterns. Areas of very high uncertainty on the RFalldata isoscape (Fig. 4d) showed lower error for EMLalldata (Fig. 4f), whereas areas with lower uncertainty in the RF approach showed higher uncertainty in the EML (Figs. 4d and 4f). The distribution of error was positively skewed for RFalldata, while for EMLalldata error had a bimodal distribution (Fig. 6). Predicted and observed bioavailable 87Sr/86Sr at EC sample sites were strongly correlated (R2 = 0.79), similar to what was observed for RFalldata.
Fig. 6.
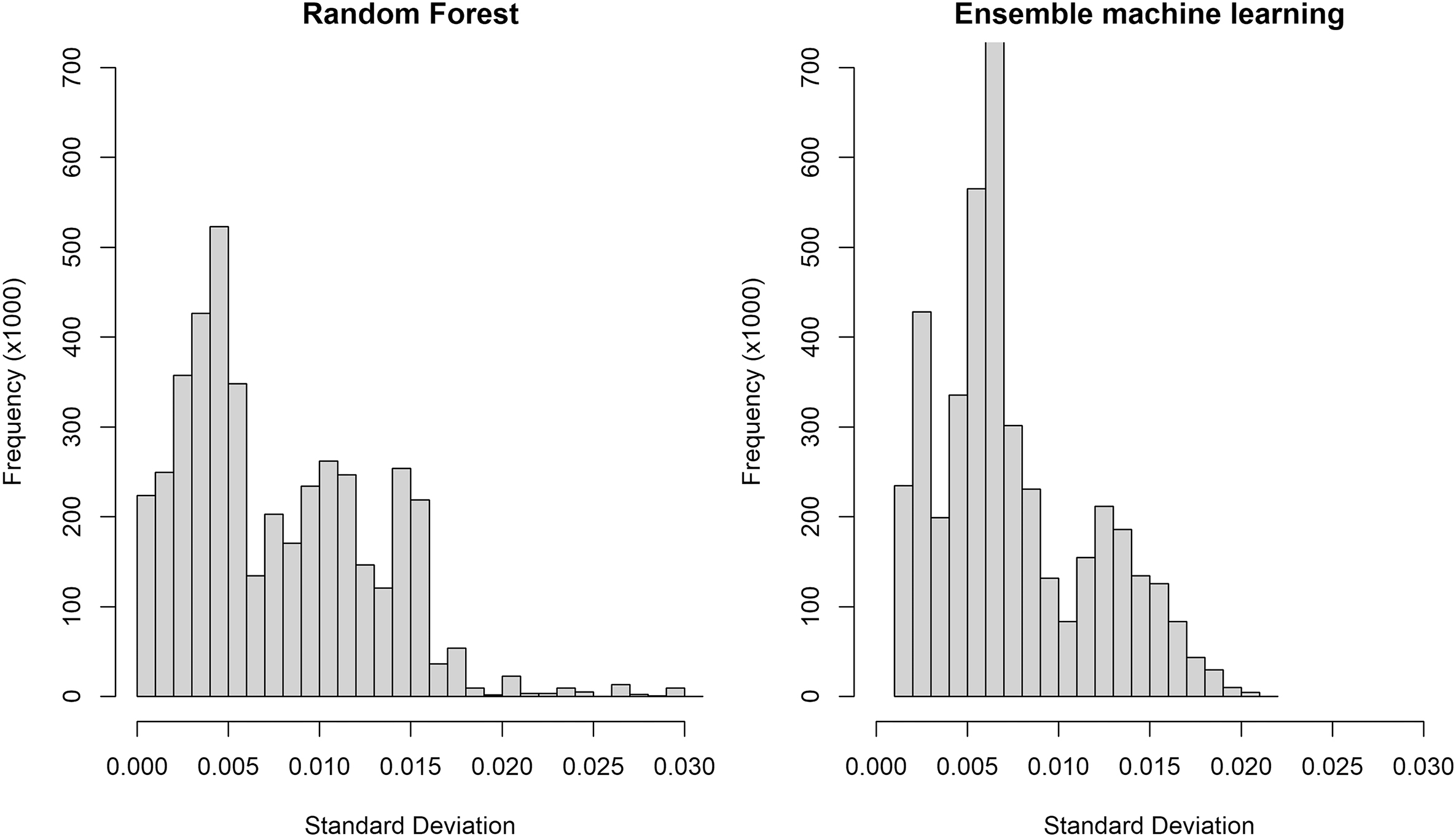
Table 2.
| Estimate | Std.Error | t value | P value | |
|---|---|---|---|---|
| (Intercept) | 5.56 | 3.693 | 1.51 | 0.132 |
| regr.ranger | 0.80 | 0.030 | 26.60 | <0.0001 |
| regr.xgboost | −9.90 | 6.556 | −1.51 | 0.131 |
| regr.nnet | −0.06 | 0.042 | −1.46 | 0.143 |
| regr.ksvm | 0.23 | 0.036 | 6.40 | <0.0001 |
| regr.cvglmnet | 0.05 | 0.037 | 1.48 | 0.140 |
Note: The variables of the model are the five base learners used to build the model. regr.ranger: random forest; regr.xgboost: extreme gradient boosting; regr.nnet: neural network; regr.ksvm: support vector machines; regr.cvglmnet: lasso and elastic-net regularised generalised linear model.
4. Discussion
Building on the previously published global bioavailable 87Sr/86Sr isoscape (Bataille et al. 2020), we produced a 87Sr/86Sr isoscape for EC using random forest and EML approaches. We added 136 sampling sites specific to EC to the original dataset of Bataille et al. (2020), improving our ability to predict bioavailable 87Sr/86Sr for this region. Moreover, we updated the geological auxiliary variables (e.g., age, lithology) using baseline lithological information for Québec with more recent and high-resolution data. Regardless of the modelling approach (RF or EML), these updates improved the spatial prediction bioavailable 87Sr/86Sr across our study area with higher 87Sr/86Sr ratios in the northern geological provinces of EC, in agreement with the 87Sr/86Sr ratios observed in our plant samples and in the literature.
The addition of EC sampling sites to the original global isoscape database (Bataille et al. 2020) increased the predicted bioavailable 87Sr/86Sr ratios of the old, highly radiogenic Canadian craton. Conversely, bioavailable 87Sr/86Sr ratios in the southern provinces (i.e., Grenville Province, the St. Lawrence platform, and the Appalachian orogeny) remained similar to predictions from Bataille et al. (2020). These provinces are dominated by carbonate and siliciclastic sedimentary units and metamorphic units, which are tectonically related to coeval North American and European units (Hartmann and Moosdorf 2012). As the global bioavailable 87Sr/86Sr isoscape from Bataille et al. (2020) was mostly trained on data-rich regions from the USA and Europe with similar lithology and climate as in southern EC, the southern EC regions show accurate bioavailable 87Sr/86Sr predictions. The addition of new bioavailable 87Sr/86Sr data from the southern EC regions only marginally improved bioavailable predictions because the machine learning model is already well informed for locations with similar lithology and environment (Bataille et al. 2020). Conversely, the Bataille et al. (2020) model performed poorly for northern EC due to the lack of bioavailable 87Sr/86Sr training data for highly radiogenic cratonic regions (i.e., less than 2% of the sites from the original database had a bioavailable 87Sr/86Sr above 0.730). In this case, even a limited number of new sites, such as the sites we added from the Superior Province, strongly helped to inform the model and drastically changed the predictions and uncertainty estimates for EC. While local predictions had a better fit with the locally calibrated bioavailable 87Sr/86Sr, the model performance only slightly improved because adding 136 sampling sites to a database of more than 4000 sampling sites globally (∼3% addition) only had a marginal effect on the overall model performance. The spatial uncertainty estimates increased significantly in northern Canada for the locally calibrated model relative to the global model because the collected bioavailable 87Sr/86Sr in northern EC showed a large range of ratios. This suggests that the machine learning model is not yet sufficiently informed in those regions and would require higher sampling density to reduce the uncertainty of the predictions across this old and lithologically heterogeneous region of the Canadian craton despite a relatively good accuracy. Similar results were found in other data-poor regions, where adding few samples improved the local model accuracy (Bataille et al. 2020; Wang et al. 2023).
The need for additional bioavailable 87Sr/86Sr data for EC is supported by the local cross-validation exercise (Fig. 5). If using the entire dataset to train a local model, this type of local cross-validation is an important step to assess the true performance of the bioavailable 87Sr/86Sr isoscape locally. We note that RMSE slowly decreased when incorporating 10%–70% EC samples into the RF training dataset (Fig. 5) then dropped for 80% and 90% EC samples. While this RMSE was lower than when no local EC samples were integrated into RF models (RMSE = 0.0112), it was much higher, even when using 90% of the EC dataset, than the RMSE found for RFalldata (model RMSE = 0.0035) and the associated local EC samples (EC data RMSE = 0.0043). This higher RMSE is not surprising because northern EC is a highly radiogenic region with heterogeneous geology and more variable bioavailable 87Sr/86Sr than many other regions of the world. This is visible in the much higher spatial uncertainty in this region (Fig. 4). Additionally, only a small portion (≈25%) of our EC dataset includes samples from northern, radiogenic EC. A large proportion of those local EC samples need to be included to see an increase in the performance of the RF model (Fig. 5). The fact that RF for 10%–90% of EC samples incorporated samples selected randomly (i.e., RF built on different datasets) increases the variability in model performance, while for 0% and 100%, for which the datasets do not change, the low variability in the RMSE estimates is solely attributable to the internal randomness of the RF algorithm. These results suggest that with every additional training sample, the model is still learning, indicating that more samples might be required to appropriately train this geologically heterogeneous cratonic area.
Variables retained to train the RF with and without EC data (RFalldata and RFnoEC, respectively) were almost identical and were consistent with Bataille et al. (2020). Differences occurred on low-ranked predictors according to the variable importance index, likely reflecting the stochasticity of the RF approach (Breiman 2001) The importance of the dust deposition variable has potentially been artificially amplified by the spatial distribution of the samples from the dataset: old and highly radiogenic regions sampled (EC, Scandinavia, Madagascar) are exposed to extremely low dust deposition. Regions heavily exposed to dust deposition are poorly represented in the dataset, and an additional sampling effort in these regions would help to better assess the importance of the dust variable. Trends observed on partial dependence were consistent between RFnoEC and RFalldata and Bataille et al. (2020). As expected, bioavailable 87Sr/86Sr increased with 87Sr/86Sr from bedrock and with bedrock age (Bataille et al. 2014, 2020). A recent application of the RF in New Zealand (Kramer et al. 2022), but using a smaller bioavailable 87Sr/86Sr dataset than Bataille et al. (2020), showed different trends in some predictors (e.g., salt deposition, mean annual temperature). Such differences are likely to reflect geographic biases and do not have clear mechanistic significance. They underline the need to build local bioavailable 87Sr/86Sr isoscapes using global bioavailable datasets instead of training them with small regional datasets. This strategy will avoid overfitting relationships between bioavailable 87Sr/86Sr and predictors yielding the most generalisable model beyond the training points.
The n-fold cross-validation of the EMLalldata demonstrated similar performance to the typical RF framework. The EMLalldata model was largely dominated by RF (highest absolute t value, Table 2) and, to a lesser extent, by support vector machine, but other learners were not as influential. Overall, the use of multiple learners did not improve the model accuracy but enhanced the robustness of the resulting model due to spatial cross-validation (Hengl et al. 2021). As expected from the dominance of RF in the EML predictions, the EMLalldata isoscape showed patterns very similar to RFalldata (Fig. 4). The main difference between spatial interpolation EML and RF-based modelling appeared when comparing the spatially explicit uncertainty maps with the EML isoscape displaying a more pronounced bimodal distribution, less positively skewed, with two normal modes (Fig. 6) reflecting the high heterogeneity of the old craton in the north and the lower variability of the younger sedimentary units in the south. The EML isoscape algorithm incorporates spatial dependencies and uses an unbiased spatial cross-validation approach, leading to more robust uncertainty estimates (Hengl et al. 2021). However, this approach is best applied to local or regional bioavailable 87Sr/86Sr predictions as computational time becomes prohibitive for larger datasets with thousands of bioavailable 87Sr/86Sr ratios (Hengl et al. 2021). Additionally, the EML framework lacks direct interpretability (i.e., there is an absence of partial dependence plots or importance of predictors), so we still recommend using the RF-based modelling approach as a basis to select predictors and discuss bioavailable 87Sr/86Sr patterns before using the EML to produce the most accurate and unbiased 87Sr/86Sr isoscape.
Further increasing the accuracy of the isoscape would require extensive sampling across EC. Intensive sampling is difficult to achieve in remote areas such as the Superior and Churchill Provinces, although collaboration with First Nations and Inuit communities and research networks conducting remote field studies may help overcome this problem. However, increasing the number of sampling sites may not be sufficient to reduce the variability in very old felsic rock units to levels similar to that of younger units. Old felsic rocks are heterogeneous with minerals having vastly different 87Sr/86Sr and weathering at different rates (Clow et al. 1997), leading to high intra-site variability (Bataille et al. 2020). Another limit of our isoscape relates to the limited number of sites across EC relative to other regions of the world. The original global dataset has a geographic bias toward the USA and Europe (Bataille et al. 2020). The link between a given predictor and the predicted 87Sr/86Sr in the global dataset may therefore not be representative of local processes, such as those observed for climate and atmospheric deposition variables (Bataille et al. 2020; Kramer et al. 2022). To refine local prediction and increase accuracy, ongoing development of the RF approach should seek to account for the spatial distribution of the sampling sites (Hengl et al. 2021; Møller et al. 2020) and to integrate local variation by weighting prediction according to the distance to the sampling sites (Sekulić et al. 2020) or to local RF (Georganos et al. 2021). However, these approaches require a relatively good spatial distribution of samples, which was not the case for our EC sampling sites. Using RF to extrapolate 87Sr/86Sr in regions with little or no data, especially in old radiogenic areas, is less reliable than in data-rich regions, and predictions should be made with caution (Bataille et al. 2020). However, we are confident in our isoscape as the predicted bioavailable 87Sr/86Sr showed good agreement with the sampling sites and are consistent with the 87Sr/86Sr from the region published in other studies (Britton 2010; Stevenson et al. 2015; Vinciguerra et al. 2016; Dargent et al. 2023).
In recent years, several provenance studies generated bioavailable 87Sr/86Sr for food and animals from EC that support the predictions of our models. For example, in the north, the 87Sr/86Sr predictions are consistent with caribou (Rangifer tarandus) 87Sr/86Sr enamel profiles from two migratory herds, the Rivière-aux-Feuilles herd and the Rivière-George herd, ranging across Québec and Labrador (Britton 2010). The range of the 87Sr/86Sr observed in enamel in the study (0.718-0.738, Britton 2010) falls within the range of the bioavailable 87Sr/86Sr observed on their annual ranges covering the Superior, Churchill, Nain Provinces, and northeast Grenville Province (Le Corre et al. 2020). Moreover, 87Sr/86Sr variations in the teeth reflect known seasonal movements, particularly for the Rivière-aux-Feuilles herd (Le Corre et al. 2020), with increasing 87Sr/86Sr values along the third molar mirroring the movement from the winter range (Le Corre et al. 2023), which encompasses the southern metamorphic region of the Superior Province in Québec, to the highly radiogenic region of the summer range in the north-west Ungava Peninsula. In the south, the isoscape is consistent with the 87Sr/86Sr measured in eastern spruce budworm moths (Dargent et al. 2023) with low ratios found along the coast of New Brunswick, Prince Edward Island, and Newfoundland (0.711-0.713), and higher ratios on the northern and southern banks of St. Lawrence River and in Gaspésie (0.715-0.718). Consistencies can also be found with data from cheese (Stevenson et al. 2015) and wine (Vinciguerra et al. 2016) studies in southern Québec.
With its unique, predictable, and stable patterns on the landscape, bioavailable 87Sr/86Sr could become a key monitoring and management tool for EC natural resources. With this developed isoscape, we have established a basis to use 87Sr/86Sr provenancing across the EC region. The EC landscape, with its broad socio-economic and biological resources, is essential to EC's economy and wealth; it also shelters great biodiversity and is the ancestral home of many Inuit and First Nations communities. Climate change and human activities are rapidly degrading EC's ecosystems and food systems (Downing and Cuerrier 2011; Foster et al. 2022). Many native species are endangered, while others (e.g., insect pests, invasive species) have seen population outbreaks costing billions of dollars annually (Logan et al. 2003). Changes in mobility are one of the key drivers of population demographics for animal species (Berger 2004; Robinson et al. 2009). Quantifying these changes through time is thus key to developing species-specific strategies to either limit (e.g., pests) or favour (e.g., endangered species) population growth. However, monitoring animal mobility is often difficult for species with large populations, species with short lifespans (e.g., insects), and species living in remote habitats (e.g., caribou). Moreover, the practical requirement for historic data that go beyond primary observations and records necessitates the development of proxy methodologies focused on historic, archaeological, or palaeontological materials. Afterall, many population drivers operate over decadal timescales requiring long-term population baselines to refine management strategies (Lindenmayer et al. 2022). Strontium isotopes could become particularly relevant to manage the biodiversity and resources of the huge EC landscape, especially for remote areas. For example, limiting the spread of eastern spruce budworm outbreaks (the most destructive insect pest of the EC boreal forest) requires the monitoring of the long-distance dispersal and connectivity of eastern spruce budworm moths, to intervene early in zones vulnerable to future outbreaks. EC would benefit from improved tools to control and mitigate those outbreaks, and the combination of hydrogen and strontium isotopes has shown promise as a way to monitor their dispersal and connectivity across the impacted regions (Dargent et al. 2023). Another example of potential use of strontium isotopes in EC is to trace the migration and range of eastern migratory caribou (Gigleux et al. 2019; Miller et al. 2023), a species central to subsistence for many Inuit and First Nations communities. With habitat alteration, human impact, climate change, and increased predation associated with the rapid development of EC, the eastern Canadian caribou population has shown a 90% decline over the last decades. For many sub-populations, demographic collapse was preceded by a change in range and mobility habits in response to human disturbances (Festa-Bianchet et al. 2011). Conserving this species will require delineating protected areas that would accommodate for range variations to allow the species to respond to future disturbances. The strontium isotope composition of caribou teeth preserves a record of the individual's mobility (Britton et al. 2009; Gigleux et al. 2019; Le Corre et al. 2023). Historical collections of caribou skeletal remains are available in museums and can be used to better delineate the long-term historical range and seasonal mobility of herds, and to help identify suitable protection areas for those endangered populations. The strontium isoscape for EC developed here therefore has immediate applicability to inform EC conservation and pest management policies for the selected species but is potentially transferable to other species (e.g., invasive species, migratory birds) with short- and long-term benefits for managing EC's historic and natural resources and biodiversity.
5. Conclusion
The wide range of 87Sr/86Sr predicted across EC make the isoscape developed here a formidable tool for provenance studies. Accounting for spatial autocorrelation between sampling sites, the EML approach showed similar model performances and predictions compare to the RF but with more robust and tighter spatial error, potentially allowing for more refined geographic assignment analyses. This approach is however better suited for local isoscapes, as it requires more computational power and may be difficult to apply at a continental scale or global scale. The isoscape we generated is the first attempt to specifically map bioavailable 87Sr/86Sr across EC. The creation of a baseline bioavailable 87Sr/86Sr isoscape that includes baseline bioavailable data from regional environmental samples is crucial for provenance studies, and our isoscape will contribute to improving studies that investigate the origin of samples or track the mobility of past and present individuals. The revised 87Sr/86Sr published here from northern Québec and Labrador are particularly valuable for the study of near- and distant-past interactions between Indigenous communities and caribou populations (Gigleux et al. 2019), a central but threatened subsistence resource. The study of the ecology of past northern species through time (e.g., Miller et al. 2022, 2023) could help us to better understand the consequences of global changes on modern northern megafauna. Our study also contributes to a broader understanding of the bioavailable 87Sr/86Sr landscape in North America, which has been documented for the USA (Bataille and Bowen 2012; Bataille et al. 2020; Funck et al. 2021), and more generally to our understanding of 87Sr/86Sr distribution in regions of old craton where data are lacking (Bataille et al. 2020). In this study, we relied on a limited number of sampling locations in EC; however, the addition of new sites across EC in future studies will serve to improve the isoscape by achieving a more representative sampling distribution, which will in turn allow the use of more complex modelling approaches (Sekulić et al. 2020; Georganos et al. 2021). However, remote areas, such as the northern part of EC, often require costly logistics and will necessitate the gathering of a broader environmental dataset from multiple sources. Data sharing, through open access repositories such as Isobank (Pauli et al. 2017), is thus an essential step in isotope mapping to produce reliable and accurate isoscapes at the local and broader scales.
Acknowledgment
We thank our colleagues from Caribou Ungava field team (J. Hénault-Richard, R. Guindon) for the plant collection, and A. Kenney and Dr. R. Lam (CREAIT, Memorial University) for assistance in the preparation and analysis of plant samples, as well as our colleagues who helped collect needles at NRCan CFS (K. Perrault, E. Owens, R. Johns, G. Leclair, J. Fidgen, and E. Moise) and in the provinces (in particular, P. Therrien and J.-J. Bertrand (Quebec MFFP), D. Lavigne and T. Rideout (Newfoundland FIA), A. McGill (Nova Scotia DNRR)).
References
Alt K.W., Knipper C., Peters D., Müller W., Maurer A.-F., Kollig I., et al. 2014. Lombards on the move—an integrative study of the migration period cemetery at Szólád, Hungary. PLoS ONE, 9: e110793.
Baffi C., Trincherini P.R. 2016. Food traceability using the 87Sr/86Sr isotopic ratio mass spectrometry. European Food Research and Technology, 242: 1411–1439.
Balmino G., Vales N., Bonvalot S., Briais A. 2012. Spherical harmonic modelling to ultra-high degree of Bouguer and isostatic anomalies. Journal of Geodesy, 86: 499–520.
Bataille C.P., Bowen G.J. 2012. Mapping 87Sr/86Sr variations in bedrock and water for large scale provenance studies. Chemical Geology, 304–305: 39–52.
Bataille C.P., Brennan S.R., Hartmann J., Moosdorf N., Wooller M.J., Bowen G.J. 2014. A geostatistical framework for predicting variations in strontium concentrations and isotope ratios in Alaskan rivers. Chemical Geology, 389: 1–15.
Bataille C.P., Crowley B.E., Wooller M.J., Bowen G.J. 2020. Advances in global bioavailable strontium isoscapes. Palaeogeography, Palaeoclimatology, Palaeoecology, 555: 109849.
Bataille C.P., Jaouen K., Milano S., Trost M., Steinbrenner S., Crubézy É., et al. 2021. Triple sulfur–oxygen–strontium isotopes probabilistic geographic assignment of archaeological remains using a novel sulfur isoscape of western Europe. PLoS ONE, 16: e0250383.
Bataille C.P., von Holstein I.C.C., Laffoon J.E., Willmes M., Liu X.-M., Davies G.R. 2018. A bioavailable strontium isoscape for Western Europe: A machine learning approach. PLoS ONE, 13: e0197386.
Beard B., Johnson C. 2000. Strontium isotope composition of skeletal material can determine the birth place and geographic mobility of humans and animals. Journal of Forensic Sciences, 45: 1049–1061.
Bentley R.A. 2006. Strontium isotopes from the earth to the archaeological skeleton: a review. Journal of Archaeological Method and Theory, 13: 135–187.
Berger J. 2004. The last mile: how to sustain long-distance migration in mammals. Conservation Biology, 18: 320–331.
Börker J., Hartmann J., Amann T., Romero-Mujalli G. 2018. Terrestrial sediments of the Earth: development of a Global Unconsolidated sediments Map database (GUM). Geochemistry, Geophysics, Geosystems, Geochemistry, Geophysics, Geosystems, 19: 997–1024.
Breiman L. 2001. Random forests. Machine learning, 45: 5–32.
Britton K. 2010. Multi-isotope analysis and the reconstruction of prey species palaeomigrations and palaeoecology. Ph.D. thesis, Durham University, Durham, UK.
Britton K., Le Corre M. 2022. Reindeer migrations at Pincevent: strontium and oxygen isotope evidence of seasonal biogeography of a key subsistence species. In Opération Archéologique Programmée Du Site de Pincevent (La Grande Paroisse, Seine-et-Marne)—Site N° 77210 AP, Autorisation Triennale 2020-2022 /Rapport Final 2022. Edited by O. Bignon-Lau. pp. 161–174.
Britton K., Grimes V., Dau J., Richards M.P. 2009. Reconstructing faunal migrations using intra-tooth sampling and strontium and oxygen isotope analyses: a case study of modern caribou (Rangifer tarandus granti). Journal of Archaeological Science, 36:1163–1172,
Britton K., Grimes V., Niven L., Steele T.E., McPherron S., Soressi M., et al. 2011. Strontium isotope evidence for migration in late Pleistocene Rangifer: implications for Neanderthal hunting strategies at the Middle Palaeolithic site of Jonzac, France. Journal of Human Evolution 61:176–85.
Britton K., Jimenez E.L., Le Corre M., Pederzani S., Daujeard C., Jaouen K., et al. 2023. Multi-isotope zooarchaeological investigations at Abri du Maras: The paleoecological and paleoenvironmental context of Neanderthal subsistence strategies in the Rhône Valley during MIS 3. Journal of Human Evolution, 174: 103292.
Britton K., Le Corre M., Willmes M., Moffat I., Grun R., Mannino, et al. 2020. Sampling plants and malacofauna in 87Sr/86Sr bioavailability studies: implications for isoscape mapping and reconstructing of past mobility patterns. Frontiers in Ecology and Evolution, 8: 579473.
Capo R.C., Stewart B.W., Chadwick O.A. 1998. Strontium isotopes as tracers of ecosystem processes: theory and methods. Geoderma, 82: 197–225.
Charlier B.L.A., Ginibre C., Morgan D., Nowell G.M., Pearson D.G., Davidson J.P., et al. 2006. Methods for the microsampling and high-precision analysis of strontium and rubidium isotopes at single crystal scale for petrological and geochronological applications. Chemical Geology, 232: 114–133.
Clow D.W., Mast M.A., Bullen T.D., Turk J.T. 1997. Strontium 87/strontium 86 as a tracer of mineral weathering reactions and calcium sources in an Alpine/Subalpine Watershed, Loch Vale, Colorado. Water Resources Research, 33: 1335–1351.
Copeland S.R., Sponheimer M., Le Roux P.J., Grimes V., Lee-Thorp J.A., De Ruiter D.J., et al. 2008. Strontium isotope ratios (87Sr/86Sr) of tooth enamel: a comparison of solution and laser ablation multicollector inductively coupled plasma mass spectrometry methods. Rapid Communications in Mass Spectrometry, 22: 3187–3194.
Coutu A.N., Whitelaw G., le Roux P., Sealy J. 2016. Earliest evidence for the ivory trade in Southern Africa: isotopic and ZooMS analysis of seventh–tenth century ad ivory from KwaZulu–Natal. African Archaeological Review, 33: 411–435.
Crowley B.E., Bataille C.P., Haak B.A., Sommer K.M. 2021. Identifying nesting grounds for juvenile migratory birds with dual isotope: an initial test using North American raptors. Ecosphere, 12: e03765.
Crowley B.E., Miller J.H., Bataille C.P. 2017. Strontium isotopes (87Sr/86Sr) in terrestrial ecological and palaeoecological research: empirical efforts and recent advances in continental-scale models. Biological Reviews, 92: 43–59.
Cutler D.R., Edwards T.C., Beard K.H., Cutler A., Hess K.T., Gibson J., et al. 2007. Random forests for classification in ecology. Ecology, 88: 2783–2792.
Czére O., Lawson J.A., Müldner G., Evans J., Boyle A., Britton K. 2022. The bodies in the ‘Bog’: a multi-isotope investigation of individual life-histories at an unusual 6th/7th AD century group burial from a roman latrine at Cramond, Scotland. Archaeological and Anthropological Sciences, 14: 67.
Dahl S.G., Allain P., Marie P.J., Mauras Y., Boivin G., Ammann, et al. 2001. Incorporation and distribution of strontium in bone. Bone, 28: 446–453.
Dargent F., Candau J.-N., Studens K., Perrault K.H., Reich M.S., Bataille C.P. 2023. Characterizing eastern spruce budworm's large-scale dispersal events through flight behavior and stable isotope analyses. Frontiers in Ecology and Evolution, 11: 10.3389.
Deniel C., Pin C. 2001. Single-stage method for the simultaneous isolation of lead and strontium from silicate samples for isotopic measurements. Analytica Chimica Acta, 426: 95–103.
Downing A., Cuerrier A. 2011. A synthesis of the impacts of climate change on the First Nations and Inuit of Canada. Indian Journal of Traditional Knowledge, 10: 57–70.
El Meknassi S., Dera G., Cardone T., De Rafélis M., Brahmi C., Chavagnac V. 2018. Sr isotope ratios of modern carbonate shells: good and bad news for chemostratigraphy. Geology, 46: 1003–1006.
Evans J.A., Montgomery J., Wildman G., Boulton N. 2010. Spatial variations in biosphere 87Sr/86Sr in Britain. Journal of the Geological Society, 167: 1–4.
Fauberteau A.E., Chartrand M.M.G., Hu L., St-Jean G., Bataille C.P. 2021. Investigating a cold case using high-resolution multi-isotope profiles in human hair. Forensic Chemistry, 22: 100300.
Festa-Bianchet M., Ray J.C., Boutin S., Cote S., Gunn A. 2011.Conservation of caribou (Rangifer tarandus) in Canada: an uncertain future. Canadian Journal of Zoology, 89: 419–434.
Font L., Nowell G.M., Pearson D.G., Ottley C.J., Willis S.G. 2007. Sr isotope analysis of bird feathers by TIMS: a tool to trace bird migration paths and breeding sites. Journal of Analytical Atomic Spectrometry, 22: 513.
Foster A.C., Wang J.A., Frost G. V, Davidson S.J., Hoy E., Turner K.W., et al. 2022. Disturbances in North American boreal forest and Arctic tundra: impacts, interactions, and responses. Environmental Research Letters, 17: 113001.
Funck J., Bataille C., Rasic J., Wooller M. 2021. A bio-available strontium isoscape for eastern Beringia: a tool for tracking landscape use of Pleistocene megafauna. Journal of Quaternary Science, 36: 76–90.
Gauthier G., Bêty J., Cadieux M.-C., Legagneux P., Doiron M., Chevallier, et al. 2013. Long-term monitoring at multiple trophic levels suggests heterogeneity in responses to climate change in the Canadian Arctic tundra. Philosophical Transactions of the Royal Society B: Biological Sciences, 368: 20120482.
Genuer R., Poggi J.-M., Tuleau-Malot C. 2015. VSURF: an R Package for variable selection using random forests. The R Journal, 7: 19–33.
Georganos S., Grippa T., Niang Gadiaga A., Linard C., Lennert M., Vanhuysse S., et al. 2021. Geographical random forests: a spatial extension of the random forest algorithm to address spatial heterogeneity in remote sensing and population modelling. Geocarto International, 36: 121–136.
Gigleux C., Grimes V., Tütken T., Knecht R., Britton K. 2019. Reconstructing caribou seasonal biogeography in Little Ice Age (Late Holocene) Western Alaska using intra-tooth strontium and oxygen isotope analysis. Journal of Archaeological Science: Reports, 23: 1043–1054.
Guibourdenche L., Stevenson R., Pedneault K., Poirier A., Widory D. 2020. Characterizing nutrient pathways in Quebec (Canada) vineyards: insight from stable and radiogenic strontium isotopes. Chemical Geology, 532: 119375.
Guiserix D., Albalat E., Ueckermann H., Davechand P., Iaccheri L.M., Bybee G., et al. 2022. Simultaneous analysis of stable and radiogenic strontium isotopes in reference materials, plants and modern tooth enamel. Chemical Geology, 606: 121000.
Hartmann J., Moosdorf N. 2012. The new global lithological map database GLiM: a representation of rock properties at the Earth surface. Geochemistry, Geophysics, Geosystems, 13: 13.
Hengl T. 2023. landmap: automated spatial prediction using ensemble machine learning[online]. Available from https://github.com/envirometrix/landmap/ [accessed 27 August 2024].
Hengl T., Miller M.A.E., Križan J., Shepherd K.D., Sila A., Kilibarda, et al. 2021. African soil properties and nutrients mapped at 30 m spatial resolution using two-scale ensemble machine learning. Scientific Reports, 11: 6130.
Hijmans R.J., Cameron S.E., Parra J.L., Jones P.G., Jarvis A. 2005. Very high resolution interpolated climate surfaces for global land areas. International Journal of Climatology, 25: 1965–1978.
Holt E., Evans J.A., Madgwick R. 2021. Strontium (87Sr/86Sr) mapping: a critical review of methods and approaches. Earth-Science Reviews, 216: 103593.
Jarvis A., Reuter A., Nelson A., Guevara E. 2008. Hole-filled SRTM for the globe Version 4, available from the CGIAR-CSI SRTM 90 m Database. CGIAR CSI Consort Spat Inf [online]. Available from http://srtm.csi.cgiar.org [accessed 27 August 2024].
Johns R.C., Bowden J.J., Carleton D.R., Cooke B.J., Edwards S., Emilson, et al. 2019. A conceptual framework for the spruce budworm early intervention strategy: can outbreaks be stopped? Forests, 10: 910.
Johnson C.M., Fridrich C.J. 1990. Non-monotonic chemical and O, Sr, Nd, and Pb isotope zonations and heterogeneity in the mafic- to silicic-composition magma chamber of the Grizzly Peak Tuff, Colorado. Contributions to Mineralogy and Petrology, 105: 677–690.
Kamenov G.D., Curtis J.H. 2017. Using carbon, oxygen, strontium, and lead isotopes in modern human teeth for forensic investigations: a critical overview based on data from Bulgaria. Journal of Forensic Sciences, 62: 1452–1459.
Kramer R.T., Kinaston R.L., Holder P.W., Armstrong K.F., King C.L., Sipple, et al. 2022. A bioavailable strontium (87Sr/86Sr) isoscape for Aotearoa New Zealand: implications for food forensics and biosecurity. PLoS ONE, 17: e0264458.
Latifovic R., Zhu Z.-L., Cihlar J., Giri C., Olthof I. 2004. Land cover mapping of North and Central America—Global Land Cover 2000. Remote Sensing of Environment, 89: 116–127.
Le Corre M., Dussault C., Côté S.D. 2020. Where to spend the winter? The role of intraspecific competition and climate in determining the selection of wintering areas by migratory caribou. Oikos, 129: 512–525.
Le Corre M., Grimes V., Lam R., Britton K. 2023. Comparison between strip sampling and laser ablation methods to infer seasonal movements from intra-tooth strontium isotopes profiles in migratory caribou. Scientific Reports, 13: 3621.
Lindenmayer D.B., Lavery T., Scheele B.C. 2022. Why we need to invest in large-scale, long-term monitoring programs in Landscape Ecology and Conservation Biology. Current Landscape Ecology Reports, 7: 137–146.
Logan J.A., Régnière J., Powell J.A. 2003. Assessing the impacts of global warming on forest pest dynamics. Frontiers in Ecology and the Environment, 1: 130–137.
Lugli F., Cipriani A., Bruno L., Ronchetti F., Cavazzuti C., Benazzi S. 2022. A strontium isoscape of Italy for provenance studies. Chemical Geology, 587: 120624.
MacLean D.A. 2019. Protection strategy against spruce budworm. Forests, 10: 1137.
Madgwick R., Lamb A.L., Sloane H., Nederbragt A.J., Albarella U., Pearson M.P., et al. 2019a. Multi-isotope analysis reveals that feasts in the Stonehenge environs and across Wessex drew people and animals from throughout Britain. Science Advances, 5: eaau6078.
Madgwick R., Lewis J., Grimes V., Guest P. 2019b. On the hoof: exploring the supply of animals to the Roman legionary fortress at Caerleon using strontium (87Sr /86Sr) isotope analysis. Archaeological and Anthropological Sciences, 11: 223–235.
Mahowald N.M., Muhs D.R., Levis S., Rasch P.J., Yoshioka M., Zender C.S., et al. 2006. Change in atmospheric mineral aerosols in response to climate: last glacial period, preindustrial, modern, and doubled carbon dioxide climates. Journal of Geophysical Research: Atmospheres, 111: D1020.
Maurer A.-F., Galer S.J.G., Knipper C., Beierlein L., Nunn E. V., Peters D., et al. 2012. Bioavailable 87Sr/86Sr in different environmental samples—effects of anthropogenic contamination and implications for isoscapes in past migration studies. Science of The Total Environment, 433: 216–229.
Miller J.H., Crowley B.E., Bataille C.P., Wald E.J., Kelly A., Gaetano, et al. 2021. Historical landscape use of migratory caribou: new insights from old antlers. Frontiers in Ecology and Evolution, 8: 590837.
Miller J.H., Fisher D.C., Crowley B.E., Secord R., Konomi B.A. 2022. Male mastodon landscape use changed with maturation (Late Pleistocene, North America). Proceedings of the National Academy of Sciences, 119.
Miller J.H., Wald E.J., Druckenmiller P. 2023. Shed female caribou antlers extend records of calving activity on the Arctic National Wildlife Refuge by millennia. Frontiers in Ecology and Evolution, 10.
Ministère des Ressources naturelles et des Forêts (MRNF). 2021a. Base de données Géologie du socle [online]. Available from https://sigeom.mines.gouv.qc.ca/signet/classes/I1102_aLaCarte [accessed 27 August 2024].
Ministère des Ressources naturelles et des Forêts (MRNF). 2021b. Base de données géochimique [online]. Available from https://sigeom.mines.gouv.qc.ca/signet/classes/I1102_aLaCarte [accessed 27 August 2024].
Møller A.B., Beucher A.M., Pouladi N., Greve M.H. 2020. Oblique geographic coordinates as covariates for digital soil mapping. SOIL, 6: 269–289.
Mooney W.D., Laske G., Masters T.G. 1998. CRUST 5.1: a global crustal model at 5° × 5°. Journal of Geophysical Research: Solid Earth, 103: 727–747.
Nguyen T.H., Watmough S.A., Dang D.H. 2023. Evaluating the use of Ca/Sr and 87Sr/86Sr ratios to track Ca sources in sugar maple in Ontario. Canadian Journal of Forest Research, 53: 772–782.
Pauli J.N., Newsome S.D., Cook J.A., Harrod C., Steffan S.A., Baker C.J.O., et al. 2017. Why we need a centralized repository for isotopic data. Proceedings of the National Academy of Sciences, 114: 2997–3001.
Pfeiffer S., Williamson R.F., Newton J., le Roux P., Forrest C., Lesage L. 2020. Population movements of the Huron–Wendat viewed through strontium isotope analysis. Journal of Archaeological Science: Reports, 33: 102466.
Poggio L., de Sousa L.M., Batjes N.H., Heuvelink G.B.M., Kempen B., Ribeiro E., et al. 2021. SoilGrids 2.0: producing soil information for the globe with quantified spatial uncertainty. SOIL, 7: 217–240.
R Core Team. 2023. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria.
Reich M.S., Flockhart D.T.T., Norris D.R., Hu L., Bataille C.P. 2021. Continuous-surface geographic assignment of migratory animals using strontium isotopes: a case study with monarch butterflies. Methods in Ecology and Evolution, 12: 2445–2457.
Robinson R., Crick H., Learmonth J., Maclean I., Thomas C., Bairlein F., et al. 2009. Travelling through a warming world: climate change and migratory species. Endangered Species Research, 7: 87–99.
Sekulić A., Kilibarda M., Heuvelink G.B.M., Nikolić M., Bajat B. 2020. Random forest spatial interpolation. Remote Sensing, 12: 1687.
Serna A., Bataille C.P., Prates L., Mange E., le Roux P., Salazar-García D.C. 2023. Small-scale mobility fostering the interaction networks of Patagonian (Argentina) hunter-gatherers during the Late Holocene: perspectives from strontium isotopes and exotic items. PLoS ONE, 18: e0281089.
Shaw H., Montgomery J., Redfern R., Gowland R., Evans J. 2016. Identifying migrants in Roman London using lead and strontium stable isotopes. Journal of Archaeological Science, 66: 57–68.
Shilts W.W., Ayslworth J.M., Kaszycki C.A., Klassen R.A. 1987. Canadian Shield. In Geomorphic Systems of North America. Edited by W.L. Graf. Geological Society of America, Boulder, Colorado, USA. pp. 119–163.
Snoeck C., Ryan S., Pouncett J., Pellegrini M., Claeys P., Wainwright A.N., et al. 2020. Towards a biologically available strontium isotope baseline for Ireland. Science of the Total Environment, 712: 136248.
Stevenson R., Desrochers S., Hélie J.-F. 2015. Stable and radiogenic isotopes as indicators of agri-food provenance: insights from artisanal cheeses from Quebec, Canada. International Dairy Journal, 49: 37–45.
Strobl C., Boulesteix A.-L., Kneib T., Augustin T., Zeileis A. 2008. Conditional variable importance for random forests. BMC Bioinformatics [Electronic Resource], 9: 307.
Sugden D.E. 1978. Glacial erosion by the Laurentide Ice Sheet. Journal of Glaciology, 20: 367–391.
Thériault R., Beauséjour S. 2012. Geological map of Québec, DV2012-07. Ministère des Ressources naturelles et des Forêts, Québec.
Vautour G., Poirier A., Widory D. 2015. Tracking mobility using human hair: What can we learn from lead and strontium isotopes? Science & Justice, 55: 63–71.
Vet R., Artz R.S., Carou S., Shaw M., Ro C.-U., Aas W., et al. 2014. A global assessment of precipitation chemistry and deposition of sulfur, nitrogen, sea salt, base cations, organic acids, acidity and pH, and phosphorus. Atmospheric Environment, 93: 3–100.
Vinciguerra V., Stevenson R., Pedneault K., Poirier A., Hélie J.-F., Widory D. 2016. Strontium isotope characterization of wines from Quebec, Canada. Food Chemistry, 210: 121–128.
Vors L.S., Boyce M.S. 2009. Global declines of caribou and reindeer. Global Change Biology, 15: 2626–2633.
Wadoux A.M.J.-C., Minasny B., McBratney A.B. 2020. Machine learning for digital soil mapping: applications, challenges and suggested solutions. Earth-Science Reviews, 210: 103359.
Wang X., Bocksberger G., Lautenschläger T., Finckh M., Meller P., O'Malley G.E., et al. 2023. A bioavailable strontium isoscape of Angola with implications for the archaeology of the transatlantic slave trade. Journal of Archaeological Science, 154: 105775.
Willmes M., Bataille C.P., James H.F., Moffat I., McMorrow L., Kinsley L., et al. 2018. Mapping of bioavailable strontium isotope ratios in France for archaeological provenance studies. Applied Geochemistry, 90: 75–86.
Willmes M., Glessner J.J.G., Carleton S.A., Gerrity P.C., Hobbs J.A. 2016. 87Sr/86Sr isotope ratio analysis by laser ablation MC-ICP-MS in scales, spines, and fin rays as a nonlethal alternative to otoliths for reconstructing fish life history. Canadian Journal of Fisheries and Aquatic Sciences, 73: 1852–1860.
Wooller M.J., Bataille C., Druckenmiller P., Erickson G.M., Groves P., Haubenstock N., et al. 2021. Lifetime mobility of an Arctic woolly mammoth. Science, 373: 806–808.
Wunder M.B. 2012. Determining geographic patterns of migration and dispersal using stable isotopes in keratins. Journal of Mammalogy, 93: 360–367.
Zhang C., Ma Y. 2012. Ensemble machine learning: methods and applications. Springer, Boston.
Zomer R.J., Trabucco A., Bossio D.A., Verchot L. V. 2008. Climate change mitigation: a spatial analysis of global land suitability for clean development mechanism afforestation and reforestation. Agriculture, Ecosystems & Environment, 126: 67–80.
Supplementary material
Supplementary Material 1 (DOCX / 14 KB).
- Download
- 13.33 KB
Supplementary Material 2 (XLSX / 750 KB).
- Download
- 749.33 KB
Supplementary Material 3 (DOCX / 3.08 MB).
- Download
- 3.08 MB
Supplementary Material 4 (R / 19 KB).
- Download
- 18.45 KB
Supplementary Material 5 (R / 47 KB).
- Download
- 46.14 KB
Information & Authors
Information
Published In

FACETS
Volume 10 • 2025
Pages: 1 - 17
Editor: Christoph E. Geiss
History
Received: 10 September 2024
Accepted: 25 October 2024
Version of record online: 12 February 2025
Copyright
© 2025 The Authors. This work is licensed under a Creative Commons Attribution 4.0 International License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Data Availability Statement
Bioavailable 87Sr/86Sr dataset generated and analysed during this study as well as R scripts are provided in full within the published article and its Supplementary material. Updated Bedrock's 87Sr/86Sr prediction maps and geological age maps as well as 87Sr/86Sr isoscapes are available in the Dryad (https://doi.org/10.5061/dryad.9zw3r22qf).
Key Words
Sections
Subjects
Plain Language Summary
A baseline strontium isoscape for provenance studies in Eastern Canada
Authors
Author Contributions
Conceptualization: MLC, JW, CPB, KB
Data curation: FD, SDC, MSR, JC, MM, BH, KB
Formal analysis: MLC, CPB
Investigation: MLC
Methodology: MLC, FD, VG, MSR, JC, MM, BH, CPB, KB
Supervision: JC, CPB, KB
Writing – original draft: MLC
Writing – review & editing: MLC, FD, VG, SDC, MSR, JC, MM, BH, CPB, KB
Competing Interests
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Funding Information
Natural Resources Canada: PLP-2019-284
Philip Leverhulme Prize: PLP-2019-284
Leverhulme Trust: RPG-2017-410
UK Research and Innovation: EP/Y023641/1
This research, including salary support for MLC, sample collection (Ungava), and isotope analysis (Memorial), was funded by The Leverhulme Trust (ref: RPG-2017-410 to KB). Further support was provided to KB during the production of this manuscript by an ERC-selected award funded by UK Research and Innovation (ref: EP/Y023641/1) and a Philip Leverhulme Prize (ref: PLP-2019-284 to KB). JC and FD received funding for the needle analysis through a Healthy Forest Partnership Early Intervention Strategy against Spruce Budworm Phase II Contribution Program awarded to the Invasive Species Centre by Natural Resources Canada. CPB received funding for this project from the NRCan Early Intervention Strategy 3.0 Small Scale Research project 2023.
Metrics & Citations
Metrics
Other Metrics
Citations
Cite As
Mael Le Corre, Felipe Dargent, Vaughan Grimes, Joshua Wright, Steeve D. Côté, Megan S. Reich, Jean-Noël Candau, Marrissa Miller, Brent Holmes, Clement P. Bataille, and Kate Britton. 2025. An ensemble machine learning bioavailable strontium isoscape for Eastern Canada. FACETS.
10: 1-17.
https://doi.org/10.1139/facets-2024-0180
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
There are no citations for this item