Superworm (Coleoptera: Tenebrionidae, Zophobas morio) degradation of UV-pretreated expanded polystyrene
Abstract
A laboratory-based study was conducted to investigate the consumption and degradation of expanded polystyrene (EPS) by superworms (darkling beetle larvae, Zophobas morio; heterotypic synonym Zophobas atratus). Superworms fed on one of three diets (wheat bran control, EPS blocks, or EPS S-shaped chips) following a 60-second pretreatment with UV radiation or no UV radiation exposure. Loss-of-mass measurements were conducted every week for 4 weeks. Nutrients (nitrogen and phosphorus) and EPS degradation products in frass were determined as well as the superworm gut microbiome composition. The average loss of mass after 28 days was 34.7% for EPS blocks and 25.6% for S-shaped chips. Small quantities of nitrogen and phosphorus were found in the frass of superworms that fed on the EPS diet, but they were lower than those from wheat bran-fed superworms. Twenty-one EPS degradation products were identified in the frass of superworms on an EPS diet, including short-chain carboxylic acids, polyols, amino acids, metabolites of amino acids, and phosphoric acids. Pseudomonas aeruginosa (polystyrene degrader) and Stenotrophomonas sp. (nitrogen fixer and phosphorus solubilization promoter) were identified from the guts of superworms that fed on an EPS diet. Overall, superworms have the potential to deal with plastic waste degradation problems.
Graphical Abstract

Introduction
Global plastic production between 2016 and 2020 exceeded 300 million tonnes annually (Plastics Europe 2021). In Canada, plastic production is a large industry that contributes $35 billion to the national economy and employs over 100 000 people. However, more than 3 million tonnes of plastic waste are disposed of in Canada every year. Despite having extensive recycling programs, only 9% of plastic materials are recycled in Canada, with a large proportion ending up in landfills or the natural environment (Environment and Climate Change Canada 2023). Moreover, ∼80%–90% of the solid waste found in marine environments is plastics of different sizes (Environment and Climate Change Canada 2023). Plastic disposal has a significant impact on human and ecological health, either directly or indirectly (Ali et al. 2021b). For example, toxic microparticles and nanoparticles are generated when polystyrene is degraded in the natural environment by either exposure to the UV radiation of the sun or physically by wind and water (Andrady 2011; Gewert et al. 2015). These micro/nano particles may induce inflammation in tissues and organs as well as hemolysis (destruction of red blood cells) (Hwang et al. 2020).
The most widely used plastics include polyethylene (PE), polypropylene (PP), polyvinyl chloride (PVC), polyethylene terephthalate (PET), and polystyrene (PS) (Andrady 2011). PS is a synthetic aromatic polymer made from styrene monomers with the molecular formula of (C8H8)n. The four main types of PS include general purpose PS (GPPS), high impact PS (HIPS), polystyrene foam (PF), and expanded polystyrene (EPS) foam, often referred to as Styrofoam (American Chemistry Council 2015; Ho et al. 2018).
PS disposal is challenging because it does not break down easily and tends to persist in the environment. In general, one of the main degradation mechanisms of synthetic polymers is through enzymatic hydrolysis (Ali et al. 2021a, 2021b). Some synthetic plastics, such as polyurethane (PU), have a C–O backbone that makes them more susceptible to enzymatic degradation. However, PS consists of a C–C backbone, which is generally difficult to break down via hydrolysis (Krueger et al. 2015; Zhang et al. 2022). In the natural environment, plastic degradation starts with photodegradation mainly via exposure to the UV radiation of the sun.
Since landfill disposal of PS is not a long-term sustainable solution, alternative options are needed to reduce PS quantities that end up in landfills and the natural environment. Recent studies have reported various invertebrate organisms that could enzymatically degrade PS. These include yellow mealworms, Tenebrio molitor L. (Yang et al. 2015a, 2015b; Brandon et al. 2018; Yang et al. 2018a, 2018b), and superworms, such as Zophobas atratus (Peng et al. 2019, 2020).
It has been reported that the microorganisms present in the gut of some insect larvae contribute to the degradation of PS (Brandon et al. 2018). For example, removal of gut bacteria via antibiotic treatment reduced biodegradation efficiency in superworms (Zophobas atratus) (Yang et al. 2020). Zophobas atratus is a heterotypic synonym of Zophobas morio. A variety of bacteria contribute to PS biodegradation in the guts of superworms. For example, Sphingobacterium spp. and Dysgonomonas spp. were found in the gut of PS-fed superworms (Luo et al. 2021). Pseudomonas spp. from superworms have also been shown to degrade PS (Kim et al. 2020). However, little information exists on the conditions that can enhance bacterial degradation, as well as the nature of degradation products, and whether these products could be used as fertilizers.
Several studies have investigated various methods to enhance degradation of PS. For example, a study has compared enzymes from various bacterial species for PS degradation (Mohanan et al. 2020). Others have explored the use of physical methods, such as UV radiation (Chamas et al. 2020), or chemical additives, such as starch, to improve PS degradation (Mohanan et al. 2020). However, studies on the potential use of superworms for PS degradation, as well as the characteristics of their degradation products, are scarce.
The objectives of this laboratory-based study were to evaluate nutrient (nitrogen and phosphorus) content and degradation product composition of frass and to identify the gut bacteria in superworms (Zophobas morio) following the ingestion of EPS.
Materials and methods
Experimental design and set-up
The study was conducted in a laboratory using a two-factor experimental design with four replicates per treatment to give a total of 24 experimental units. The two factors studied were as follows: Factor 1 was the type of diet fed to the superworms, which was either S-shaped EPS chips or block-type EPS or wheat bran as the control. The S-shaped EPS is typically anti-static and of lower density (3.8– 4.0 kg/m3), whereas the block-type EPS is typically higher density (14–16 kg/m3). Factor 2 was the type of pre-treatment applied to the polystyrene samples, which was either a 60-second direct exposure to UV radiation using a transilluminator (Fisher Scientific Model FTBI-614 312 nm with total UV intensity of 8000 µw/cm2) or no UV radiation. The UV treatment time of 60 s was selected because it was short enough to minimize exposure to surrounding environment, cost-effective if this could be adopted by waste management facilities, and potentially long enough to loosen bonds with the polystyrene structure and make it easier to break down. The mass of EPS used in each container was 1.44 g.
The superworms were initially fed a wheat bran diet before the experiment started and were then starved for 48 h before being fed only S-shaped EPS chips, block-type EPS, or wheat bran (control). A total of 12 superworms were placed in each 500 mL plastic beaker and covered with perforated parafilm to prevent superworms from escaping. The study was conducted at a room temperature, approximately 19–20 °C. Diet treatments were sprayed with deionized water once a day. No nutrients were added to either treatment.
Following a 28-day incubation period, EPS loss of mass was measured, and the frass of superworms was collected and analyzed. Total nitrogen and phosphorus contents of frass were determined using UV–visible spectrophotometry.
Loss of mass and survival rates
The loss of mass measurements (in grams) were conducted on a weekly basis for 4 weeks. It involved carefully removing the intact EPS blocks or S-shaped chips in each container and weighing them. The small EPS particles resulting from superworms eating the chips or blocks were not weighed as these were scrapped off using a small brush. The percent loss of mass at the end of each week was calculated by dividing the loss-of-mass of that week by the original mass of EPS in each container (i.e., 1.44 g). The loss of mass rate (in mg per day per superworm) was calculated at the end of the 28-day period for each container by dividing the total loss of mass in that container by 28 and then by the number of superworms that survived the experiment. Survival rate was determined as the total number of superworms that survived each week until 28 days of feeding on one of three diets divided by the number of superworms at the beginning of the feeding period and was expressed as a percentage.
Statistical analysis
Statistical analysis was conducted using the Statistical Package for Social Sciences (SPSS) Version 26. The Analysis of variance (ANOVA) was conducted using the general linear model procedure to assess the data of each week. The sources of variation included in ANOVA were the type of diet, the type of pre-treatment applied to the polystyrene samples, and the potential interaction between them. Statistical significance was determined when P value (ANOVA) < 0.05.
Preparation of frass samples for chemical analysis
Different sieve sizes (0.5 and 1.0 mm) were used to separate large EPS particles from the frass. The final separation was conducted by blowing air gently in a receiving pan. The frass from all four replicates per treatment were pooled prior to chemical (i.e., nitrogen, phosphorus, and degradation products) analysis.
Nitrogen analysis
Nitrogen extraction of frass was performed using modified Kjeldahl method (APHA 1999) in three steps: digestion, distillation, and titration. A precisely weighed frass sample (∼0.1 g) was placed into a Kjeldahl flask. A total of 6 mL of H2SO4 and 2.89 g of K2SO4 were added to the flask plus a small piece of copper as the catalyst to increase the reaction rate of converting organic nitrogen into ammonium and therefore reduce the reaction time. The flask was then heated up to 370 °C for 75 min.
Distillation was conducted after the solution cooled down to room temperature. A total of 100 mL of deionized water was added to the flask. The digest was made basic by adding approximately 25 mL of 45% w/v NaOH to the flask to convert ammonium (NH4+) into ammonia gas. Ammonia (NH3) was then steam distilled into a solution made of 28 mL distilled water mixed with 6 mL of 12 mol/L HCl. The solution was then titrated with standardized 1 mol/L NaOH solution to the colorless end point using phenolphthalein as indicator.
Phosphorus analysis
Phosphorus determination was conducted using the modified rapid digestion method by Pequerul et al. (1993). A total of 0.25–0.30 g of the air-dried frass sample was digested in 5 mL of 15% H2O2 and 5 mL of concentrated (15.8 mol/L) HNO3. The digest was cooled down, syringe filtered (2.5 µm), washed with 5 mL of 6 mol/L HCl, and centrifuged (269 ✕ g, 10 min). The supernatant was then diluted to 25 mL using chromatography-grade water and acidified with 5 mL of 1:1 HCl, to ensure that final pH was much lower than 7. Phosphorus content was determined in the supernatant (10 mL) using the ammonium molybdate/stannous chloride method (APHA 1999) with absorbance measured at 690 nm.
Analysis of polystyrene degradation products in frass
Five hundred microliters of 50% methanol in chromatography-grade water were added to 100 mg of frass and mixed vigorously (3 ✕ 20 s) then centrifuged (15 000 ✕ g, 15 min, 4 °C). The supernatant (200 µL) was filtered (0.46 µm) and dried under nitrogen (30 °C). Extracted compounds were derivatized with bis(trimethylsilyl)trifluoroacetamide (85 µL, 80 °C, 1 h) and analyzed by a Bruker Scion 436 TQ Gas Chromatography-Mass Spectrometer (GC-MS/MS) instrument on a HP-5MS 30 m ✕ 0.250 mm ✕ 0.25 µm column (Agilent Technologies) using an adaptation of a published method (Tsochatzis et al. 2021). Briefly, derivatized compounds (1.0 µL) were injected into the GC column at a 10:1 split ratio with an injector temperature of 300 °C. The oven temperature was held at 60 °C for 2 min, then raised to 290 °C at a rate of 10 °C/min, and held at 290 °C for 5 min. Full-scan MS analysis was performed for m/z 50–600 with transfer line temperature and source temperature both set to 290 °C. The total analysis time was 30 min. Major GC-MS peaks (>2 GCps) were extracted and their MS spectra were searched against the Human Metabolome Database (HMDB) version 5.0 using its GC-MS search tool with MS tolerance set to ±0.5 Da (Wishart et al. 2022). Only compounds with a proportion of the library spectrum’s peaks with matches > 0.80 were considered. A library match score > 0.80 means at least 80% of the MS peaks obtained from the sample were identical to those in the library spectrum i.e., compound was a “match”. Unusually derivatized (such as trimethylsilylated compounds) and physiologically irrelevant compounds (i.e., compounds not supposed to be found in the sample, such as drugs or household products) were excluded from the search.
Metabolic pathway analysis
Pathway analysis was performed on the identified HMDB hits using the MetaboAnalyst 5.0 web tool for metabolomics data analysis (Pang et al. 2022) with hypergeometric testing and relative betweenness centrality set for enrichment and topology analyses, respectively, in Drosophila melanogaster as the closest species in the web tool to Zophobas morio. Significantly altered pathways were visualized on a scatter plot with pathway impact and ‒log10p on the x- and y-axes, respectively. Pathway impact was calculated by the web tool as the sum of importance measures of each of the matched metabolites divided by the sum of the importance measures of all metabolites in each pathway.
Preparation of polystyrene film
To form polystyrene films, 1.5 g of EPS pieces were dissolved in xylene and the solution was left to dry for 24 h. The film was removed, washed with 10 mL of 100% methanol and then rinsed with deionized water (DW), and then left to dry for three more days.
Liquid carbon-free media
Liquid carbon-free medium (LCFBM) was prepared by dissolving 0.7 g of KH2PO4, 0.7 g of K2HPO4, 0.7 g of MgSO4·7H2O, 1.0 g of NH4NO3, 0.005 g of NaCl, 0.002 g of FeSO4·7H2O, 0.002 g of ZnSO4·7H2O, and 0.001 g of MnSO4·H2O in 1 L of DW. Luria-Bertani (LB) broth medium was prepared by dissolving 25 g of LB Miller broth into 1 L of DW. To make the solid version, 15 g of agarose was added to the medium. All the media were autoclaved at 121 °C for 15 min.
Extraction of gut bacteria
Bacteria were extracted from larvae that had been fed EPS for 28 days as their sole diet. The larvae were submerged in 70% ethanol for 3 min. After submersion the larvae were dissected, and three larval guts were placed in a 15 mL tube containing 3 mL of liquid carbon free media. The tubes were shaken for 15 min and the gut tissue was removed. The tubes were centrifuged (448 ✕ g, 5 min) at room temperature to obtain a pellet followed by removal of supernatant and pellet resuspension in 5 mL of LCFBM. The washing step was repeated two more times with the final pellet being resuspended in 1 mL of LCFBM.
Isolation and identification of bacteria
The collected gut cell suspension was used to inoculate liquid cultures containing 0.50 g of sterile EPS pieces or EPS film in a 250 mL Erlenmeyer flask (n = 4) with 100 mL of LCFBM. The cultures were incubated at 24 °C and 120 RPM for 10–11 days of growth; dilutions of 1/10, 1/100, and 1/1000 of the liquid cultures were plated onto LB or bovine heart infusion for colony isolation.
Colonies from the plates were selected based on colony morphology (shape, colour, and size). Each colony was added to a 5 mL culture tube with 2 mL of LB and incubated at 24 °C and 120 RPM for 24 h.
To establish that the isolated bacteria were able to grow on this media as an individual species, an inoculum from each colony was added to LCFBM containing EPS. Samples were taken at regular intervals and examined by spectrophotometer to assay for increased numbers of cells.
DNA extraction and amplification
Bacterial DNA was isolated using a Blood and Tissue kit from Qiagen following the instructions for gram-positive cells included in the kit. The concentration of DNA was determined by a nanodrop spectrophotometer. For those bacteria that displayed degradation capabilities, their 16S rRNA gene was amplified by PCR targeting the hyper-variable V4 region using the following primers:
•
forward primer 515 F (5′-GTGCCAGCMGCCGCGGTAA-3′)
•
reverse primer 806R (5′-GGACTACHVGGGTWTCTAAT-3′).
The final volume of this PCR was 50 µL, with 12.5 µL trehalose (20%), 5 µL buffer, 3 µL MgCl2 (50 mM), 1 µL dNTPs (10 mM), 2.5 µL forward primer (10 µmol/L), 2.5 µL reverse primer (10 µmol/L), 1 µL Taq polymerase (5 U/µL), and 100 ng of template DNA. PCR was performed under the following conditions at 94 °C for 2 min, 94 °C for 30 s, 56 °C for 40 s, 72 °C for 1 min, 72 °C for 10 min. The PCR products were sent to Genome Quebec for 16S rRNA gene sequencing. The returned sequences were Basic Local Alignment Search Tool (BLAST) searched in the NCBI database and compared to the observed bacteria characteristics to confirm identification.
Results
Polystyrene loss of mass and superworm survival rates
Figures 1–4 show the disintegration of the denser EPS blocks compared to the less dense and anti-static S-shaped chips following the consumption by superworms. The superworms created tunnels and holes through the blocks and chips, and in the process small EPS particles (micro and macroparticles) were generated. The ANOVA results for the loss-of-mass (%) data showed significant differences between EPS blocks and S-shaped chips for each of the four weeks. After 7 days of consumption by the superworms, the average loss-of-mass for EPS blocks was 16.0%, which was significantly higher compared to 5.5% for the S-shaped chips (F = 20.926; P value = 0.000639). After 14 days of consumption the observed average loss-of-mass of 24.4% for the EPS blocks showed an increase compared to the previous week and was significantly greater than the 11.8% for S-shaped chips (F = 16.465; P value = 0.001588). The loss-of-mass continued to increase after 21 days of consumption by superworms and that for the EPS blocks was 30.6%, which was significantly higher than the 15.3% observed for the S-shaped EPS chips (F = 22.940; P value = 0.000441). Results for the final week (i.e., 28 days after consumption of EPS by superworms) of the loss-of-mass observations also indicated a significantly higher average loss of 34.7% for the EPS blocks compared to 25.6% for the S-shaped EPS chips (F = 25.670; P value = 0.000277).
Fig. 1.

Fig. 2.

Fig. 3.

Regression analysis results for loss-of-mass (%) versus time (days) indicated that the loss-of-mass percentages generally increased over time as expected, due to more consumption of the EPS material. However, the rate of increase in the loss-of-mass percentage was significantly greater for the denser EPS blocks compared to the anti-static and less dense S-shaped EPS chips. All four regression plots shown in Fig. 5 indicated very strong linear or curvilinear relationships (R2 > 0.95 in all cases) between loss-of-mass percentage and number of days of consumption by superworms.
The average loss-of-mass rate in mg per day per superworm at the end of the 28-day period was also compared among diets and the results indicated that there was a significantly higher (F = 16.342; P = 0.002) rate of EPS loss (per day per superworm) for the EPS blocks compared to the EPS S-shaped chips. The average loss of mass rate of EPS was 0.17 mg per day per superworm for blocks and was significantly higher than the 0.10 mg per day per superworm for the S-shaped chips.
ANOVA results for the loss-of-mass (%) also indicated that there were no significant differences between UV compared to no UV treatments (P > 0.05). After 28 days of consumption by superworms the average loss-of-mass percentages for UV-treated EPS was 25.6%, which was not significantly different from 27.0% observed for the no UV treatment (F = 0.175; P value = 0.683). Exposure of EPS to UV radiation also did not have a significant effect on the EPS loss of mass rate per day per superworm (F = 0.031; P = 0.863). The UV and the no UV pre-treatments of EPS prior to consumption by superworms resulted in the same loss of mass rate of 0.14 and 0.13 mg per day per superworm, respectively.
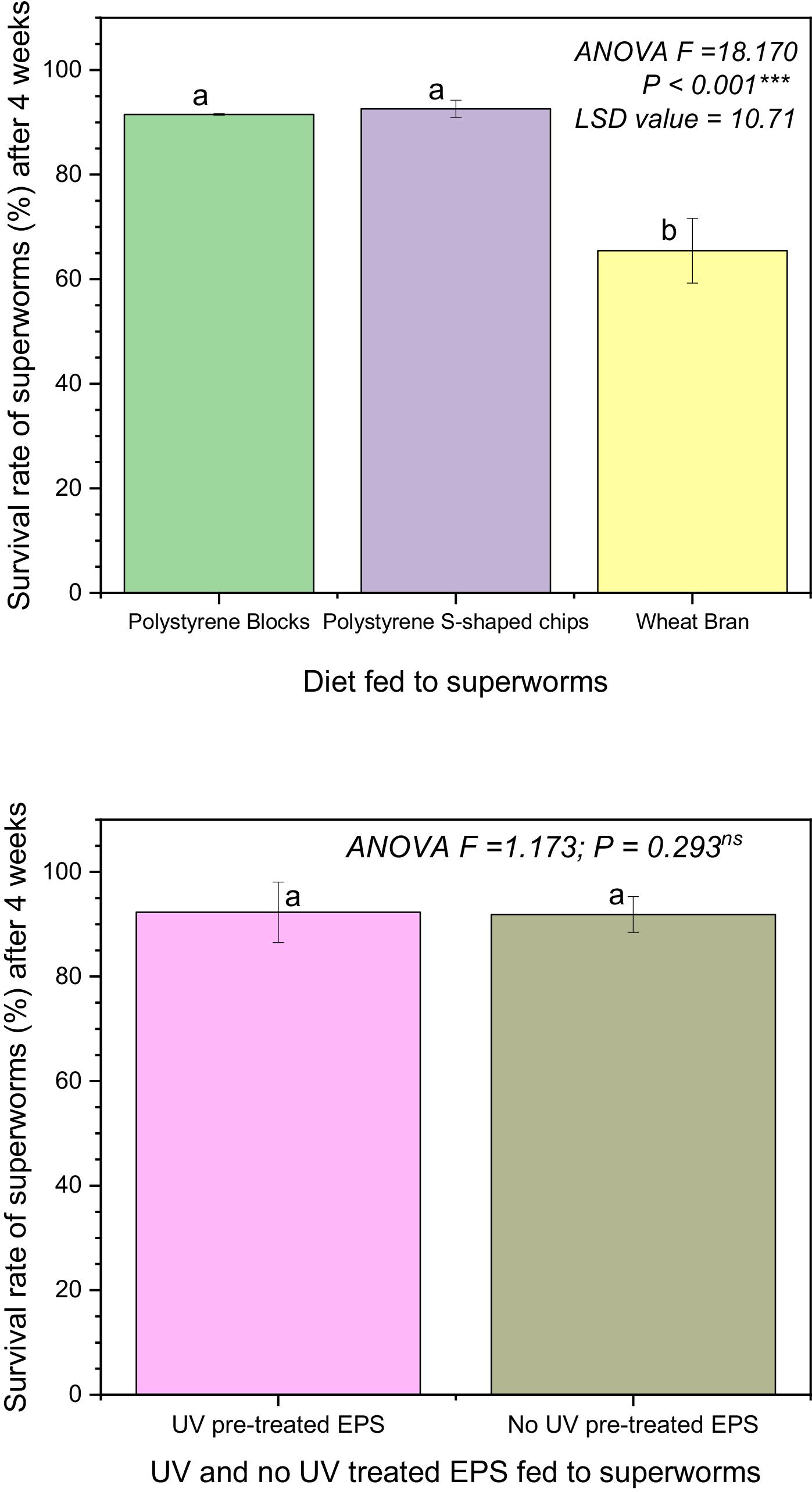
Survival rate of superworms
The results of statistical analysis showed significant differences among some diet treatments impact on survival rates (ANOVA F = 18.17; P < 0.001). Post-hoc comparison using the least significance difference test indicated that the survival rate (as %) of superworms after 4 weeks of feeding on 100% wheat bran was surprisingly much lower than that of superworms that had been fed to EPS blocks and EPS S-shaped chips (see Fig. 6). The survival rates for superworms fed to EPS blocks and EPS S-shaped chips were not significantly different from each other (P > 0.05) and they were both greater than 90%, while that under wheat bran was at 65%.
The UV exposure pre-treatment of the diet before being fed to the superworms did not affect their survival rates. On average the survival rates of superworms under UV pre-treated versus non-UV pre-treated EPS diet were also greater than 90%. Some of the superworms continued to feed on the EPS several weeks beyond the initial 4 weeks of the study, and some pupate and then turned into adult beetles that continued to feed on EPS diet (see Fig. 4).
Fig. 4.

Fig. 5.
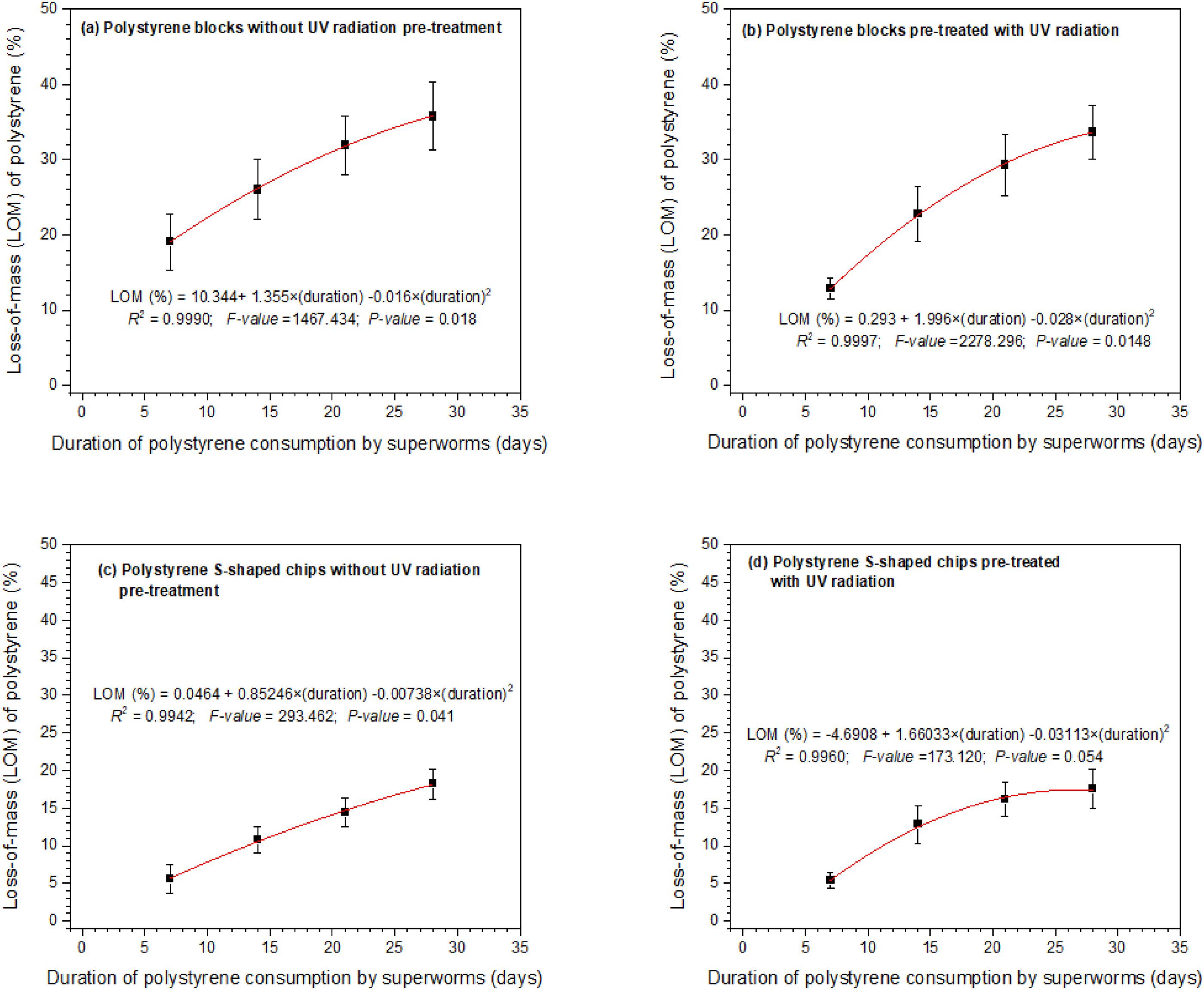
Fig. 6.
Frass phosphorous and nitrogen content
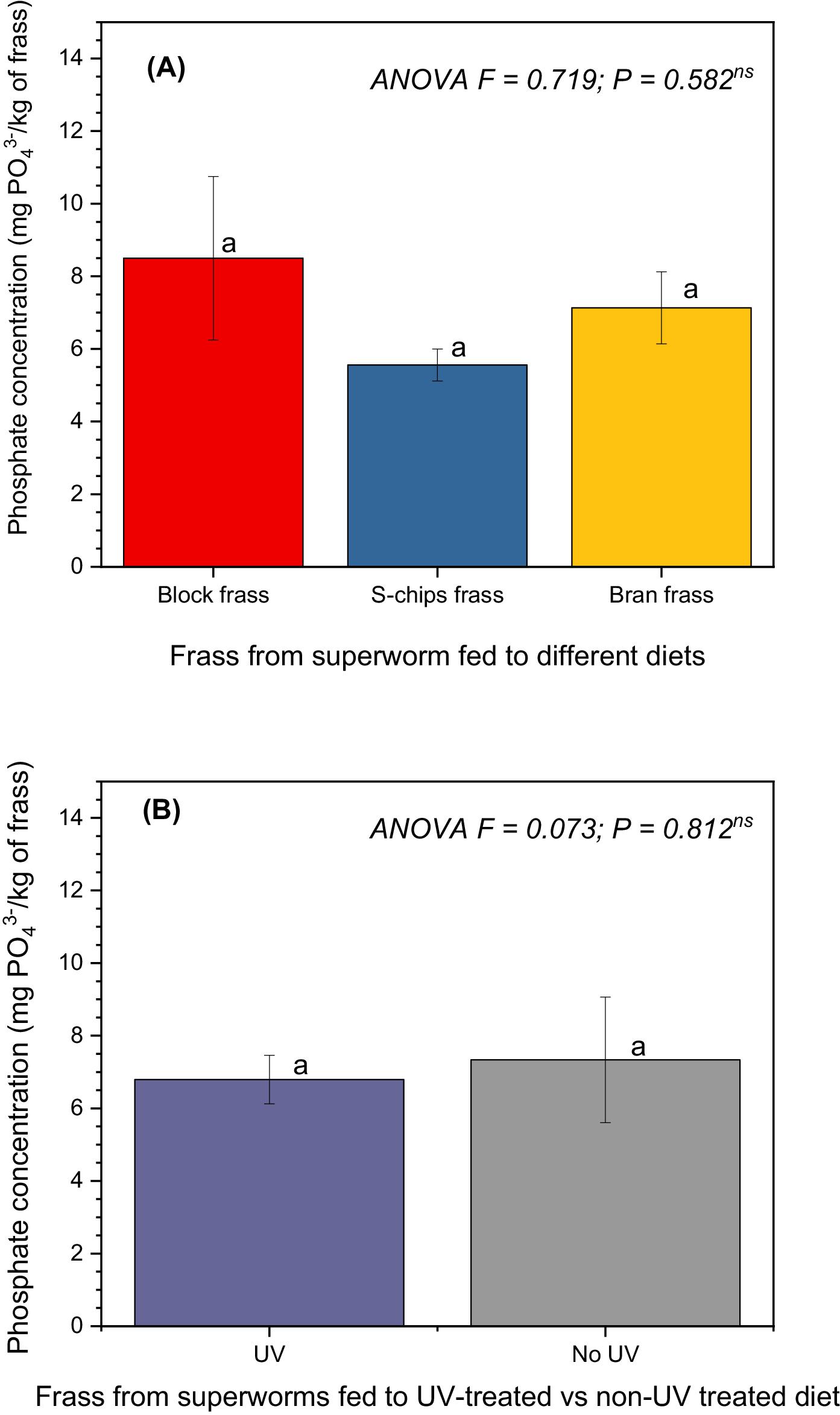
ANOVA results showed that phosphate (PO43−) content did not differ significantly (ANOVA F = 0.719; P = 0.582) in the frass of superworms that had been fed to EPS diet (blocks or S-shaped chips) and wheat bran (see Fig. 7). Similarly, there were no differences in PO43− content between frass from superworms that had been fed on UV-pretreated diet compared to non-UV pretreated diet (ANOVA F = 0.073; P = 0.812). The PO43− content in frass averaged between 5.6 and 8.5 mg/kg of frass from the S-shaped EPS chips and EPS blocks, respectively. These equate to elemental phosphorus (P) content of 1.8 and 2.8 mg/kg for the S-shaped EPS chips and EPS blocks, respectively.
Fig. 7.
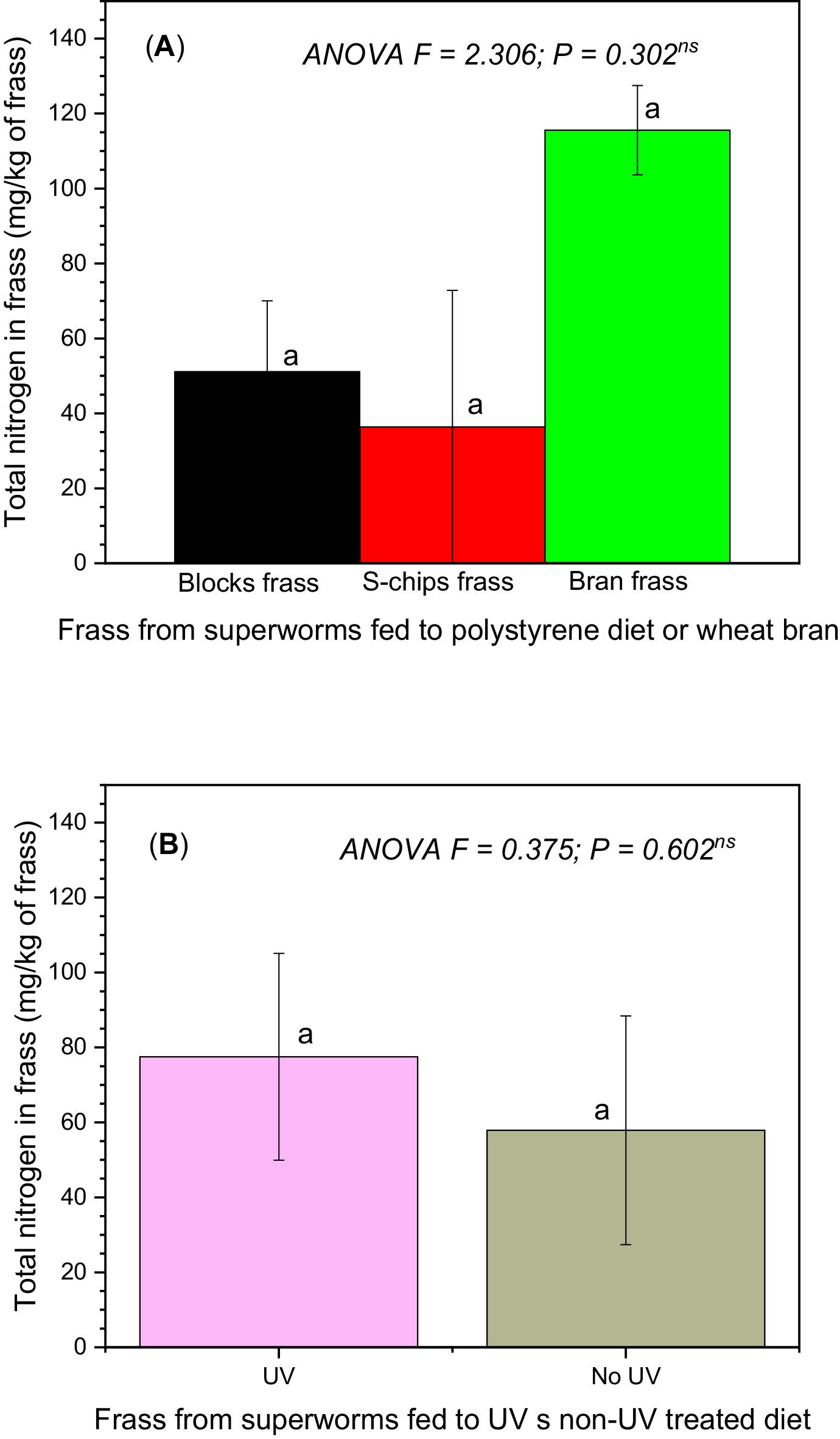
The nitrogen concentration in frass samples was not significantly different among diets (i.e., EPS blocks, EPS S-shaped chips, and wheat bran) fed to the superworms (ANOVA F = 2.306; P = 0.302) as shown in Fig. 8. Similarly, no differences occurred between nitrogen concentrations in frass samples regardless of whether the diet fed to the superworms had been pre-treated with 60 s of UV exposure or not (ANOVA F = 0.375; P = 0.602).
Fig. 8.
PS degradation products in superworm frass
Twenty-one compounds were tentatively identified by GC-MS/HMDB in the hydrophilic extract of EPS-fed superworm frass with most of these compounds being amino acids (seven compounds) and short-chain carboxylic acids (six compounds). Two polyols were identified (2-amino-2-methyl-1,3-propanediol and glycerol) as well as two diamines (putrescine and cadaverine), two phosphorous-containing compounds (phosphoric acid and pyrophosphoric acid), and two aromatic compounds (kynurenic acid and terephthalic acid). The average molecular weight of the identified compounds was 122.4 Da, with an average molecular formula of C3.8H5.3N0.9O2.9P0.1. Out of the identified compounds, two did not include any carbon atoms (phosphoric acid and pyrophosphoric acid) and two did not contain any oxygen atoms (putrescine and cadaverine). No sulfur-, halogen-, or metal-containing compounds were identified. Table 1 contains the complete list of compounds tentatively identified in EPS-fed superworm frass by GC-MS.
Table 1.
| Compound | Molecular formula | Molecular weight, Da | GC retention time, min | Matching score* |
|---|---|---|---|---|
| Alanine | C3H7NO2 | 89.1 | 7.6 | 1.00 |
| 3-Aminoisobutanoic acid | C4H9NO2 | 103.1 | 7.8 | 1.00 |
| 2-Amino-2-methyl-1,3-propanediol | C4H11NO2 | 105.1 | 2.9 | 0.86 |
| Arginine | C6H14N4O2 | 174.2 | 11.7 | 0.82 |
| Cadaverine | C5H14N2 | 102.2 | 12.3 | 1.00 |
| Glutamic acid | C5H9NO4 | 147.1 | 14.5 | 0.96 |
| Glycolic acid | C2H4O3 | 76.0 | 7.1 | 1.00 |
| Glycerol | C3H8O3 | 92.1 | 10.3 | 1.00 |
| Histidine | C6H9N3O2 | 155.2 | 17.6 | 1.00 |
| Hydroxyproline | C5H9NO3 | 133.1 | 13.5 | 1.00 |
| Kynurenic acid | C10H7NO3 | 189.2 | 18.9 | 1.00 |
| Lactic acid | C3H6O3 | 90.1 | 6.9 | 1.00 |
| Methylmalonic acid | C4H6O4 | 118.1 | 6.9 | 1.00 |
| Oxalic acid | C2H2O4 | 90.0 | 1.8 | 1.00 |
| Phosphoric acid | H3PO4 | 98.0 | 10.1 | 1.00 |
| Phthalic acid | C8H6O4 | 166.1 | 16.3 | 1.00 |
| Putrescine | C4H12N2 | 88.2 | 14.6 | 1.00 |
| Pyroglutamic acid | C5H7NO3 | 129.0 | 13.1 | 1.00 |
| Pyrophosphoric acid | H4P2O7 | 178.0 | 10.2 | 0.95 |
| Succinic acid | C4H6O4 | 118.1 | 10.5 | 1.00 |
| Valine | C5H11NO2 | 117.2 | 9.3 | 1.00 |
*
Matching score refers to the proportion of HMDB library spectrum peaks with matches.
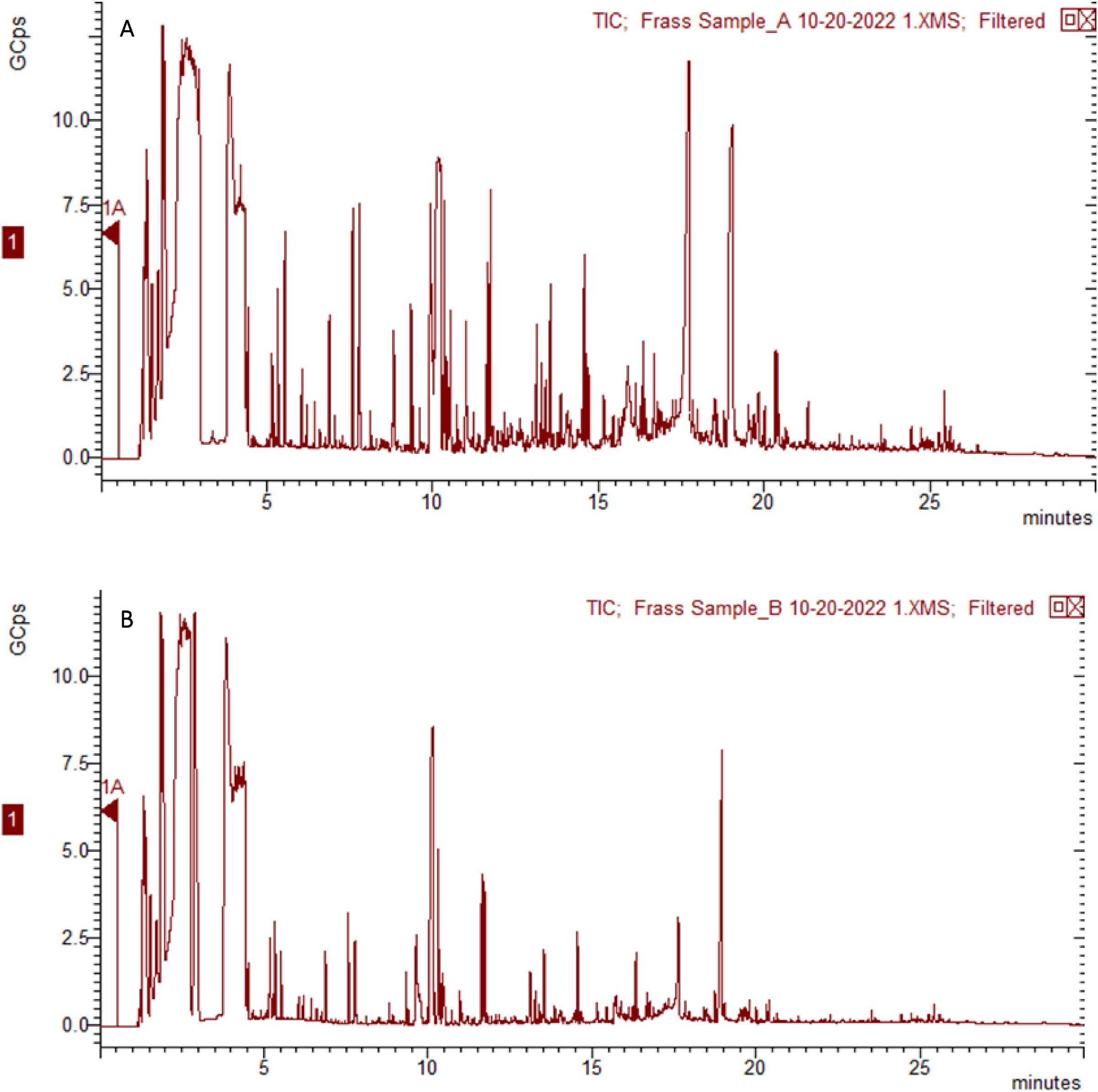
Frass samples from superworms fed on UV-treated EPS had a more complex GC-MS total ion chromatogram (TIC) and a larger average peak area in comparison with samples from superworms fed on untreated samples. Figure 9 exhibits TIC plots from two representative frass samples (UV-treated versus untreated). TICs of frass samples from superworms feeding on EPS chips were not different to those from the superworms feeding on EPS blocks.
Fig. 9.
Pathway analysis identified aminoacyl-tRNA biosynthesis being the most significantly altered pathway in the superworms fed on an EPS diet followed by glutathione metabolism, arginine and proline metabolism, alanine, aspartate and glutamate metabolism, and butanoate metabolism. Figure 10 visualizes these pathways along with other less significantly altered metabolic pathways. The results from the pathway analysis are included in Table 2.
Fig. 10.
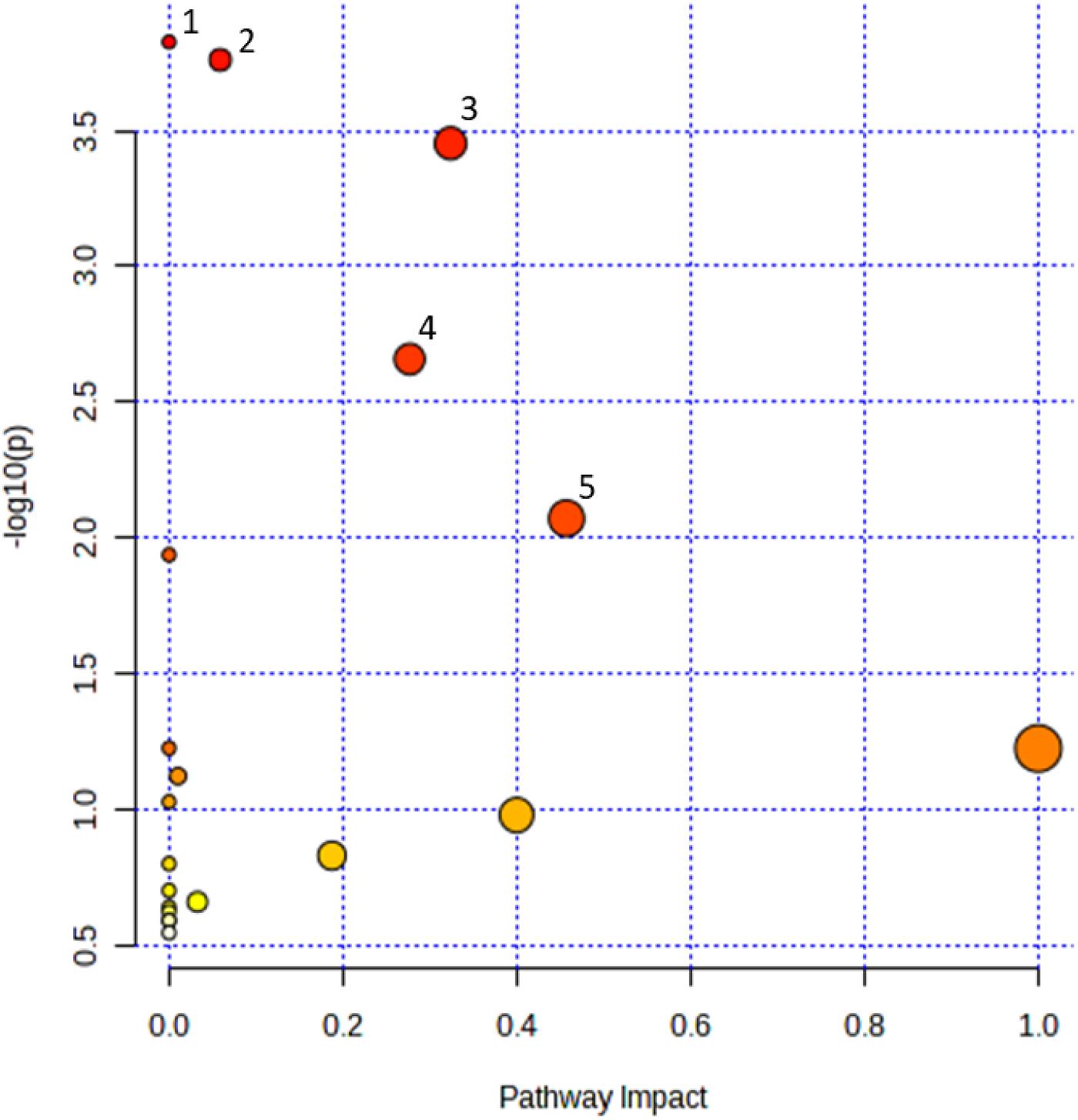
Table 2.
| Pathway name | Total metabolites in pathway | Metabolites identified in pathway | Corrected P | FDR | Impact |
|---|---|---|---|---|---|
| Aminoacyl-tRNA biosynthesis | 48 | 5 | 0.0120 | 0.0070 | 0.00 |
| Glutathione metabolism | 26 | 4 | 0.0139 | 0.0070 | 0.06 |
| Arginine and proline metabolism | 31 | 4 | 0.0278 | 0.0095 | 0.32 |
| Alanine, aspartate, and glutamate metabolism | 23 | 3 | 0.1712 | 0.0444 | 0.28 |
| Arginine biosynthesis | 12 | 2 | 0.6541 | 0.1376 | 0.46 |
| Butanoate metabolism | 14 | 2 | 0.8779 | 0.1559 | 0.00 |
| Nitrogen metabolism | 5 | 1 | 1.0000 | 0.6047 | 0.00 |
| D-Glutamine and D-glutamate metabolism | 5 | 1 | 1.0000 | 0.6047 | 1.00 |
| Valine, leucine, and isoleucine degradation | 38 | 2 | 1.0000 | 0.6806 | 0.01 |
| Valine, leucine and isoleucine biosynthesis | 8 | 1 | 1.0000 | 0.7611 | 0.00 |
| Histidine metabolism | 9 | 1 | 1.0000 | 0.7740 | 0.40 |
| Glycerolipid metabolism | 13 | 1 | 1.0000 | 0.9907 | 0.19 |
| beta-Alanine metabolism | 14 | 1 | 1.0000 | 0.9907 | 0.00 |
| Pantothenate and CoA biosynthesis | 18 | 1 | 1.0000 | 1.0000 | 0.00 |
| Citrate cycle (TCA cycle) | 20 | 1 | 1.0000 | 1.0000 | 0.03 |
| Propanoate metabolism | 21 | 1 | 1.0000 | 1.0000 | 0.00 |
| Pyruvate metabolism | 22 | 1 | 1.0000 | 1.0000 | 0.00 |
| Glyoxylate and dicarboxylate metabolism | 24 | 1 | 1.0000 | 1.0000 | 0.00 |
| Porphyrin and chlorophyll metabolism | 24 | 1 | 1.0000 | 1.0000 | 0.00 |
| Galactose metabolism | 27 | 1 | 1.0000 | 1.0000 | 0.00 |
Note: FDR represents false-discovery rate. Impact is the sum of importance measures of each of the matched metabolites divided by the sum of the importance measures of all metabolites in each pathway.
Identification of gut bacteria based on 16S rRNA gene sequencing.
The 16S amplicons were sequenced directly with the 806R primers using Sanger sequencing. The 16S rRNA generated for isolates WEP1SWBLUV20, WEP13SWBLUV20, and WEP14SWBLUV20 had 210, 223, and 188 bases, respectively. Strain identification by BLAST showed that isolate WEP1SWBLUV20 was Stenotrophomonas sp. Isolates WEP13SWBLUV20 and WEP14SWBLUV20 were confirmed by sequencing to be Pseudomonas aeruginosa. Table 3 shows the percentage similarity of the analyzed strains against representative species. All three identified strains were able to grow in LCFBM with EPS as the sole carbon source.
Table 3.
| Number of isolate | Representative species | Percent similarity |
|---|---|---|
| WEP1SWBLUV20 | Stenotrophomonas sp. strain H3-1 | 100% |
| WEP13SWBLUV20 | Pseudomonas aeruginosa strain HS337 | 99% |
| WEP14SWBLUV20 | Pseudomonas aeruginosa strain consensus data 16S rRNA gene | 100% |
Discussion and conclusions
The loss-of-mass rates per day per superworm observed in our study was within the same range reported for mealworm larvae (Tenebrio molitor L.) which ranged between 0.04 and 0.4 mg per day per mealworm observed in related studies (Brandon et al. 2018; Yang et al. 2018a; Yang et al. 2021; Palmer et al. 2022). More specifically the loss-of-mass rate for the EPS fed to mealworms over a 32-day period averaged 0.10 mg per day per mealworm (Yang et al. 2021), which was equal to what we also observed in our study for superworms fed on expanded EPS over a 28-day period. In a separate studyYang et al. (2020) observed that superworms (Zophobas atritus) consumed EPS at a rate of 0.58 mg per day per superworm, which was four times higher than consumption rate by mealworms (T. molitor). That study also reported that the superworms were able to live normally on an EPS diet for 28 days.
The decrease in the mass of EPS S-shaped chips and white blocks over the 28-day period was significant. The decrease over time was relatively consistent, regardless of whether the polystyrene had been pre-treated with 60 s of exposure to UV radiation.
The phosphorus content of 1.8–2.8 g/kg frass observed in our study is much lower than that reported for insect frass contents of up to 20 g/kg frass (Houben et al. 2020). However, in our study regardless of whether the superworm larvae fed on EPS diets (S-shaped chips and white blocks) or wheat bran, the total P content was not different, indicating that this could be perhaps the naturally low P content found in superworm frass. Although the specific details of the composition of EPS used in this study are not available, it is common for PS manufactured to have additives that contain nitrogen and phosphorus. Four additives commonly used in PS production and have significant nitrogen content include 2-(2′-hydroxy-5′-methylphenyl)benzotriazole) (Tinuvin P), bis (2, 2, 6, 6-tetramethyl-4-piperidyl) sebacate (Tinuvin 770), which are both used in PS production as UV absorbers to stabilize PS when exposed to UV radiation, N, N-ethylene-bis(tetrabromophthalimide) (Saytex BT-93) which is used as flame retardant, and a N, N’-ethylene-bis-stearamide-based synthetic wax (Acrawax) used as a lubricant (Ho et al. 2018). These four additives are used at rates of 0.2%–0.3% for Tinuvin P, 0.2%–0.5% for Tinuvin 770, up to 13% for Saytex BT-93, and 200 ppm for Acrawax, and therefore likely provided a source of nitrogen for the superworms feeding on the EPS diet. Thirteen of the 21 degradation products that we identified in frass were nitrogen-containing molecules, including an amine diol, an aromatic amine, 2 aliphatic diamines, and 7 amino acids.
Another additive used in PS production is tris(nonylphenyl) phosphite (Wytox) that contains phosphorus, and its antioxidant properties reduce the degradation of PS (Ho et al. 2018). These compounds can be used in PS production at amounts of 0.2%–0.3% for Tinuvin P and 0.2% for Wytox (Ho et al. 2018). Therefore, the phosphorus observed in our frass samples may have originated from PS additives used during its manufacture. Part of this phosphorus was found to be biotransformed into phosphoric acid and pyrophosphoric acid.
Our results showed that the pre-treatment of EPS with UV radiation exposure for 60 s did not result in significant differences in loss of mass, nitrogen content in frass, and phosphorus content in frass. However, degradation products identified from GC-MS analyses of frass showed more abundance of amino acids and short-chain carboxylic acids in frass from superworms fed on EPS diet. Pre-treatment with 60 s of UV radiation exposure resulted in high quantities of degradation products compared to non-UV treated frass samples. A study by de Castro Monsores et al. (2021) showed that UV pre-treatment of PS resulted in modification of the macromolecules which affected its mechanical properties, such as scission of molecular chains. Other studies using various organisms have confirmed that UV radiation pre-treatment can enhance degradation of plastics. For example, Esmaeili et al. (2013) reported that exposure of PE to Aspergillus niger fungal species for 126 days resulted in degradation percentages of 30% for UV treated versus 16% for non-UV treated PE. Similarly enhanced degradation of UV pre-treated polypropylene (PP) by Phanerochaete chrysosporium NCIM 1170 (F1) and Engyodontium album MTP091 (F2) fungi was also reported (Jeyakumar et al. 2013). Similar to what has been reported for plastic degradation in natural environments, the UV pre-treatment of PS initiates photo-oxidation that causes cleavage of polymer bonds resulting in shorter chain oxygen containing products that are more prone to biodegradation by microorganisms (Yousif and Haddad 2013).
Some studies have reported that frass from superworms fed to polystyrene diets had high content of amides (Gan et al. 2021). However, we did not identify any amides among the most abundant compounds in our frass samples. A study by Jiang et al. (2021) identified several alcohols and carboxylic acids in the gut and frass of superworms that had fed on PS diet, including phthalic acid, which was identified in our analysis as well. However, compared to the aforementioned study, we identified carboxylic acids with a shorter carbon chain, such as succinic acid, glycolic acid, and lactic acid.
Chemical characterization studies have reported total nitrogen contents of up to 50 g/kg frass (i.e., equal to 5% or 5000 mg/kg frass) from mealworms that had exclusively been fed to wheat bran diet (Houben et al. 2020).
Previous studies (Peng et al. 2020, 2022a, 2022b) have used gel permeation chromatography, coupled with Fourier-transform infrared spectroscopy to monitor the process of PS degradation in mealworm and superworm frass. We chose to use GC-MS as it allowed us to also characterize the degradation products.
Our results show that PS exposure has triggered several metabolic pathways in superworms. A study by Peng et al. (2022b) equally showed many metabolic pathways found in mealworm (Tenebrio molitor) gut. Among these pathways, the pathways that overlapped between the two studies are arginine and proline metabolism, D-glutamine and D-glutamate metabolism, and histidine metabolism.
No previous studies have reported the identification of amino acids in the frass of superworms.
A study found that some species, such as larval zebrafish, exhibited increased levels of some amino acids such as glutamine, beta-alanine, or some acids such as methylmalonic acid and succinic acid, while also decreasing in the levels of putrescine in the presence of PS (Wan et al. 2018).
The gut microbial community of superworms is thought to contribute to the degradation of EPS. In our study superworm gut bacteria were isolated to identify species that may be responsible for EPS degradation. The isolates were grown on media containing EPS as the sole carbon source and DNA from the isolates was extracted, then the V4 region of the 16S rRNA gene was amplified by PCR and sequenced. One of sequences analyzed by BLAST was identified to belong to Stenotrophomonas sp. The genus Stenotrophomonas is composed of at least sixteen species that have been identified from various sources (Deng et al. 2022). The main environmental reservoirs of the Stenotrophomonas spp. are plants and soil. These gram-negative bacteria are mainly non-pathogenic except for the opportunistic pathogen Stenotrophomonas maltophilia. Most species of Stenotrophomonas promote plant growth via several mechanisms including nitrogen fixation (Park et al. 2005), solubilization of phosphate, as well as protecting the plant from pathogens and promoting growth under stressing conditions (Ryan et al. 2009). Members of this genera have been found to persist in the gut of the bark beetle Dendroctonus rhizophagus throughout its life cycle (Morales-Jiménez et al. 2012).
To the best of our knowledge Stenotrophomonas sp. has not previously been identified as a component of the gut microbial community in superworm larvae. This genus of bacteria has been identified from the gut of meal worms that have been fed an exclusively EPS diet and a polyethylene diet that included bran (Lou et al. 2021). Stenotrophomonas sp. isolated from solid waste dump sites has been shown to degrade low-density polyethylene (Nadeem et al. 2021). Stenotrophomonas produces polyamines like putrescine and cadaverine that correlate with root development and could protect the plants from environmental stresses (Ulrich et al. 2021). Decarboxylation of ornithine or arginine is the first step in the synthesis of polyamines (Ulrich et al. 2021). It is interesting to note that polyamines and arginine were found among the degradation products tentatively identified in EPS-fed superworm frass Table 1.
We also identified Pseudomonas sp. as constituents of the superworm gut. A study by Kim et al. (2020) reported that of all the gut bacteria identified in superworms using DNA sequencing analysis, 35% were Pseudomonas aeruginosa. Our study identified Pseudomonas aeruginosa from superworm guts, the same bacteria species that has also been identified by other researchers in superworm guts (Kim et al. 2020; Lee et al. 2020; Li et al. 2020). Pseudomonas aeruginosa isolated from guts of superworms has been found to degrade PS at a rate of 0.1% per day, and the serine hydrolase enzyme secreted by this bacteria species was involved in PS degradation (Lee et al. 2020).
In conclusion, our study results showed significant loss-of-mass of EPS blocks and S-shaped chips after 28 days of consumption by superworms. Survival rates of superworms that relied on EPS diet alone were greater than 90% for both EPS blocks and S-shaped chips. Pre-treatment using UV exposure for 60 s did not affect loss of mass rates for EPS, or nitrogen and phosphorus content in frass. Frass from the superworms contained nitrogen and phosphorus, and therefore could potentially serve as a fertilizer. GC-MS analyses of frass showed that UV pretreatment resulted in higher peak areas and more complex degradation products that included amino acids and short-chain carboxylic acids. Gut bacteria isolated from the superworms using PCR amplification and 16S rRNA gene sequencing followed by BLAST search showed sequence identity with Pseudomonas aeruginosa and Stenotrophomonas sp. Our study findings suggest that superworms have the potential to degrade polystyrene, however further research is needed to determine whether this could be a sustainable approach to managing polystyrene.
Acknowledgements
We thank Devin Hughes, Emily Pollock, Anthony Sousa, and Marina Grintsov for their assistance with laboratory equipment maintenance, calibration and training of research assistants that worked on this project. Funding for this research came from three Concordia University of Edmonton (CUE) Internal Research Grants. Thank you to our industry collaborators, Westlock Regional Waste Management Centre (Tom Moore) and Alberta Recycling and Management Authority (Annie Lam) who were involved in discussions on initiating this phase of the Polystyrene Degradation Project (PDP).
References
Ali S.S., Elsamahy T., Al-Tohamy R., Zhu D., Mahmoud Y.A.-G., Koutra E., et al. 2021a. Plastic wastes biodegradation: mechanisms, challenges, and future prospects. Science of The Total Environment, 780: 146590.
Ali S.S., Elsamahy T., Koutra E., Kornaros M., El-Sheekh M., Abdelkarim E.A., et al. 2021b. Degradation of conventional plastic wastes in the environment: a review on current status of knowledge and future perspectives of disposal. Science of the Total Environment, 771: 144719.
American Chemistry Council. 2015. Plastics foodservice packaging group PFPG FDA task force. The safety of styrene-based polymers for food-contact use 2013. Available from https://www.plasticfoodservicefacts.com/wp-content/uploads/2017/10/Polystyrene-Report.pdf[Accessed 4 May 2023].
APHA. 1999. Standard methods for the examination of water and wastewater. 20th ed. American Public Health Association, Washington, DC.
Andrady A.L. 2011. Microplastics in the marine environment. Marine Pollution Bulletin, 62: 1596–1605.
Brandon A.M., Gao S.H., Tian R., Ning D., Yang S.S., Zhou J., et al. 2018. Biodegradation of polyethylene and plastic mixtures in mealworms (larvae of Tenebrio molitor) and effects on the gut microbiome. Environmental Science & Technology, 52(11): 6526–6533.
Chamas A., Moon H., Zheng J., Qiu Y., Tabassum T., Jang J.H., et al. 2020. Degradation rates of plastics in the environment. ACS Sustainable Chemistry & Engineering 2020, 8(9): 3494–3511.
Deng Y., Han X.F., Jiang Z.M., Yu L.Y., Li Y., Zhang Y.Q. 2022. Characterization of three Stenotrophomonas strains isolated from different ecosystems and proposal of Stenotrophomonas mori sp. nov. and Stenotrophomonas lacuserhaii sp. nov. Frontiers in Microbiology, 13(13): 1056762.
Castro Monsores K.G., da Silva A.O., de Sant’ Ana Oliveira S., Weber R.P., Filho F.P., Monteiro S.N. 2021. Influence of ultraviolet radiation on polystyrene. Journal of Materials Research and Technology, 13: 359–365.
Environment and Climate Change Canada. 2023. Plastic waste and pollution reduction. Available from https://www.canada.ca/en/environment-climate-change/services/managing-reducing-waste/reduce-plastic-waste.html (Accessed 5 May 2023).
Esmaeili A., Pourbabaee A.A., Alikhani H.A., Shabani F., Esmaeili E. 2013. Biodegradation of low-density polyethylene (LDPE) by mixed culture of Lysinibacillus xylanilyticus and Aspergillus niger in soil. PLoS ONE, 8(9): e71720.
Gan S.K.E., Phua S.X., Yeo J.Y., Heng Z.S.L., Xing Z. 2021. Method for zero-waste circular economy using worms for plastic agriculture: augmenting polystyrene consumption and plant growth. MDPI, 4(43): 1–10.
Gewert B., Plassmann M.M., MacLeod M. 2015. Pathways for degradation of plastic polymers floating in the marine environment. Environmental Science: Processes & Impacts, 17: 1513–1521.
Ho B.T., Roberts T.K., Lucas S. 2018. An overview of the biodegradation of polystyrene and modified polystyrene: the microbial approach. Critical Reviews in Biotechnology, 38(2): 308–320.
Houben D., Daoulas G., Faucon M.P., Dulaurent A.M. 2020. Potential use of mealworm frass as a fertilizer: impact on crop growth and soil properties. Scientific Reports, 10: 4659.
Hwang J., Choi D., Han S., Jung S.Y., Choi J., Hong J. 2020. Potential toxicity of polystyrene microplastic particles. Scientific Reports, 10: 7391.
Jeyakumar D., Chirsteen J., Doble M. 2013. Synergistic effects of pretreatment and blending on fungi mediated biodegradation of polypropylenes. Bioresource Technology, 148: 78–85.
Jiang S., Su T., Zhao J., Wang Z. 2021. Biodegradation of polystyrene by Tenebrio molitor, Galleria mellonella, and Zophobas atratus larvae and comparison of their degradation effects. Polymers, 13(20): 3539.
Kim H.R., Lee H.M., Yu H.C., Jeon E., Lee S., Li J.J., Kim D.H. 2020. Biodegradation of polystyrene by Pseudomonas sp. isolated from the gut of superworms (larvae of Zophobas atratus). Environmental Science & Technology, 54: 6987–6996.
Krueger M.C., Harms H., Schlosser D. 2015. Prospects for microbiological solutions to environmental pollution with plastics. Applied Microbiology and Biotechnology, 99: 8857–8874.
Lee H.M., Kim H.R., Jeon E., Yu H.C., Lee S., Li J., Kim D.H. 2020. Evaluation of the biodegradation efficiency of four various types of plastics by Pseudomonas aeruginosa isolated from the gut extract of superworms. Microorganisms, 8(9): 1341.
Li J., Kim H.R., Lee H.M., Yu H.C., Jeon E., Lee S., Kim D.H. 2020. Rapid biodegradation of polyphenylene sulfide plastic beads by Pseudomonas sp. Science of the Total Environment 720: 137616.
Luo L., Wang Y., Guo H., Yang Y., Qi N., Zhao X., et al. 2021. Biodegradation of foam plastics by Zophobas atratus larvae (Coleoptera: Tenebrionidae) associated with changes of gut digestive enzymes activities and microbiome. Chemosphere, 282: 131006.
Lou Y., Li Y., Lu B., Liu Q., Yang S., Liu B., et al. 2021. Response of the yellow mealworm (Tenebrio molitor) gut microbiome to diet shifts during polystyrene and polyethylene biodegradation. Journal of Hazardous Materials, 416: 126222.
Mohanan N., Montazer Z., Sharma P.K., Levin D.B. 2020. Microbial and enzymatic degradation of synthetic plastics. Frontiers in Microbiology, 11: 580709.
Morales-Jiménez J., Zúñiga G., Ramírez-Saad H.C., Hernández-Rodríguez C. 2012. Gut-associated bacteria throughout the life cycle of the bark beetle Dendroctonus rhizophagus Thomas and Bright (Curculionidae: Scolytinae) and their cellulolytic activities. Microbial Ecology, 64: 268–278.
Nadeem H., Alia K.B., Muneer F., Rasul I., Siddique M.H., Azeem F., Zubair M. 2021. Isolation and identification of low-density polyethylene degrading novel bacterial strains. Archives of Microbiology 203: 5417–5423.
Palmer K.J., Lauder K., Christopher K., Guerra F., Welch R., Bertuccio A.J. 2022. Biodegradation of expanded polystyrene by larval and adult stages of Tenebrio molitor with varying substrates and beddings. Environmental Processes, 9(3): 1–15.
Pang Z., Zhou G., Ewald J., Chang L., Hacariz O., Basu N., Xia J. 2022. Using MetaboAnalyst 5.0 for LC-HRMS spectra processing, multi-omics integration and covariate adjustment of global metabolomics data. Nature Protocols, 17: 1735–1761.
Park M., Kim C., Yang J., Lee H., Shin W., Kim S., Sa T. 2005. Isolation and characterization of diazotrophic growth promoting bacteria from rhizosphere of agricultural crops of Korea. Microbiological Research, 160(2): 127–133.
Pequerul A., Pérez C., Madero P., Val J., Monge E. 1993. A rapid wet digestion method for plant analysis. Vol. 53. In Optimization of plant nutrition. Developments in plant and soil sciences. Edited by M.A.C. Fragoso, M.L. Van Beusichem, A. Houwers. Springer, Dordrecht.
Peng B.Y., Sun Y., Chen Z., Chen J., Zhou X.F., Benbow M.E., et al. 2019. Biodegradation of polystyrene by dark (Tenebrio obscurus) and yellow (Tenebrio molitor) mealworms (Coleoptera: Tenebrionidae). Environmental Science & Technology, 53: 5256–5265.
Peng B.Y., Li Y., Fan R., Chen Z., Chen J., Brandon A.M., et al. 2020. Biodegradation of low-density polyethylene and polystyrene in superworms, larvae of Zophobas Atratus (Coleoptera: Tenebrionidae): broad and limited extent depolymerization. Environmental Pollution, 266: 115206.
Peng B.Y., Sun Y., Wu Z., Chen J., Shen Z., Zhou X., et al. 2022a. Biodegredation of polystyrene and low-density polyethylene by Zophobas atratus larvae: fragmentation into microplastics, gut microbiota shift, and microbial functional enzymes. Journal of Cleaner Production, 367(3): 132987.
Peng B.Y., Sun Y., Xiao S., Chen J., Zhou X., Wu W.M., Zhanh Y. 2022b. Influence of polymer size on polystyrene biodegradation in mealworms (Tenebrio molitor): responses of depolymerization pattern, gut microbiome, and metabolome to polymers with low to ultrahigh molecular weight. Environmental Science & Technology, 56(23): 17310–17320.
Plastics Europe. 2021. Plastics—the facts 2021, an analysis of European plastics production, demand, and waste data. Available from https://plasticseurope.org/knowledge-hub/plastics-the-facts-2021/.
Ryan R., Monchy S., Cardinale M., Taghavi S., Crossman L., Avison M.B., et al. 2009. The versatility and adaptation of bacteria from the genus Stenotrophomonas. Nature Reviews Microbiology, 7: 514–525.
Tsochatzis E.D., Berggreen I.E., Nørgaard J.V., Theodoridis G., Dalsgaard T.K. 2021. Biodegradation of expanded polystyrene by mealworm larvae under different feeding strategies evaluated by metabolic profiling using GC-TOF-MS. Chemosphere, 281: 130840.
Ulrich K., Kube M., Becker R., Schneck V., Ulrich A. 2021. Genomic analysis of the endophytic Stenotrophomonas strain 169 reveals features related to plant-growth promotion and stress tolerance. Frontiers in Microbiology, 12: 687463.
Wan Z., Wang C., Zhou J., Shen M., Wang X., Fu Z., Jin Y. 2018. Effects of polystyrene microplastics on the composition of the microbiome and metabolism in larval zebrafish. Chemosphere, 217: 646–658.
Wishart D.S., Guo A., Oler E., Wang F., Anjum A., Peters H., et al. 2022. HMDB 5.0: the Human Metabolome Database for 2022. Nucleic Acids Research, 50(D1): D622–D631.
Yang Y., Yang J., Wu W.M., Zhao J., Song Y.L., Gao L.C., et al. 2015a. Biodegradation and mineralization of polystyrene by plastic-eating mealworms: part 1. chemical and physical characterization and isotopic tests. Environmental Science & Technology, 49: 12080–12086.
Yang Y., Yang J., Wu W.M., Zhao J., Song J.L., Gao L.C., et al. 2015b. Biodegradation and mineralization of polystyrene by plastic-eating mealworms: part 2. Role of gut microorganisms. Environmental Science & Technology, 49: 12087–12093.
Yang S.S., Brandon A.M., Flanagan J.C., Yang J., Ning D., Cai S.Y., et al. 2018a. Biodegradation of polystyrene wastes in yellow mealworms (larvae of Tenebrio molitor Linnaeus): factors affecting biodegradation rates and the ability of polystyrene-fed larvae to complete their life cycle. Chemosphere, 191: 979–989.
Yang S.S., Wu W.M., Brandon A.M., Fan H.Q., Receveur J.P., Li Y., et al. 2018b. Ubiquity of polystyrene digestion and biodegradation within yellow mealworms, larvae of tenebrio molitor Linnaeus (Coleoptera: Tenebrionidae). Chemosphere, 212: 262–271.
Yang Y., Wang J., Xia M. 2020. Biodegradation and mineralization of polystyrene by plastic-eating superworms Zophobas atratus. Science of the Total Environment, 708(708): 135233.
Yang L., Gao J., Liu Y., Zhuang G., Peng X., Wu W.-M., Zhuang X. 2021. Biodegradation of expanded polystyrene and low-density polyethylene foams in larvae of Tenebrio molitor Linnaeus (Coleoptera: Tenebrionidae): broad versus limited extent depolymerization and microbe-dependence versus independence. Chemosphere, 262: 127818.
Yousif E., Haddad R. 2013. Photodegradation and photostabilization of polymers, especially polystyrene: review. SpringerPlus, 2: 398.
Zhang Y., Pedersen J.N., Eser B.E., Guo Z. 2022. Biodegradation of polyethylene and polystyrene: from microbial deterioration to enzyme discovery. Biotechnology Advances, 60: 107991.
Information & Authors
Information
Published In

FACETS
Volume 9 • 2024
Pages: 1 - 15
Editor: Peter G. Kevan
History
Received: 7 May 2023
Accepted: 24 January 2024
Version of record online: 27 June 2024
Copyright
© 2024 The Author(s). This work is licensed under a Creative Commons Attribution 4.0 International License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Data Availability Statement
Data will be made available upon request.
Key Words
Sections
Subjects
Plain Language Summary
Break down of plastic by superworms: analysis of their feces and gut bacteria
Authors
Author Contributions
Conceptualization: EM, DH, CB, MG
Data curation: EM, DH, SN
Formal analysis: EM, DH, SA, EP, SN, MM, MG
Funding acquisition: EM, DH, CB, MG
Investigation: EM, DH, CB, SA, EP, SN, MM, MG
Methodology: EM, DH, SA, EP, SN, MM, MG
Project administration: EM, DH
Resources: EM, DH, CB, EP, MM, MG
Supervision: EM, DH, MG
Validation: MG
Visualization: EM
Writing – original draft: EM, DH, SA, MG
Writing – review & editing: EM, DH, CB, SN
Competing Interests
The authors declare there are no competing interests.
Funding Information
Concordia University of Edmonton: CRG-SEED-2105-02, CRG-SEED-2110-01
Metrics & Citations
Metrics
Other Metrics
Citations
Cite As
Emmanuel Mapfumo, Deborah Hemmerling, Cecilia Bukutu, Sachin Acharya, Emma Paterson, Seth Nobert, Megan MacElheren, and Makan Golizeh. 2024. Superworm (Coleoptera: Tenebrionidae, Zophobas morio) degradation of UV-pretreated expanded polystyrene. FACETS.
9: 1-15.
https://doi.org/10.1139/facets-2023-0075
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
Cited by
1. Assessing the impact of weathered polystyrene collected from the marine environment on oxidative stress responses in Zophobas morio larvae: A preliminary study