The effect of nutrient transport downstream on food web stability in an experimental freshwater meta-ecosystem
Abstract
Recent spatial nutrient transport theory suggests that accumulation of nutrients downstream in riverine systems can amplify the magnitude of phytoplankton and zooplankton blooms and/or lead to competitive replacement of phytoplankton by less edible species, such as cyanobacteria. We used an experimentally controlled three-node network of freshwater mesocosms with green algae, cyanobacteria, and Daphnia magna to test these hypotheses. Nutrients and detritus accumulated significantly downstream, reaching maximum values in the terminal nodes, resulting in small increases in abundance for green algae and D. magna populations. Stability analysis from the Lotka–Volterra competition model for green algae and cyanobacteria did not provide evidence of complete competitive exclusion, but cyanobacteria projected to equilibrate at densities 50% higher than those of green algae in all nodes. Our results support the theoretical prediction that unidirectional flow in riverine systems contributes to accumulation of nutrients downstream and increased heterotrophic bacterial activity, but these changes were of insufficient magnitude to produce variance- or mean-driven destabilization of food web relationships downstream in our experimental system.
Introduction
The escalating demand for global crop production has increased inorganic nitrogen and phosphorus fertilizer usage from 10 and 4.4 Tg year−1, respectively, to 102 and 20 Tg year−1, from 1961 to 2021 (FAO 2022). As the world’s population approaches the projected 9.7 billion milestone (United Nations 2022), nitrogen and phosphorus fertilizers required to support the expanding agricultural production is expected to increase up to 158 and 27 Tg year−1 by 2050 (FAO 2018; Mogollón et al. 2018; Nedelciu et al. 2020). While synthetic fertilizers provide essential nutrients for high crop yield, 55% of global nitrogen and 40% of phosphorus inputs are lost to the atmosphere through volatilization, or to ground and surface waters through leaching and runoff (Bouwman et al. 2009; Yadav et al. 2017; Ros et al. 2020; Zhang et al. 2021; Zou et al. 2022). In certain regions of world such as Asia and Oceania, this loss can be as high as 70% due to variation in the quality and quantity of fertilizers applied, crop types, soil characteristics, and climatic conditions (Bouwman et al. 2009; Zhang et al. 2021).
Much of this nutrient flux is due to fertilizer run-off entering different water bodies, such as groundwater, streams, lakes, and coastal environments. This advective process will likely continue to escalate, potentially disrupting normal patterns of nutrient cycling and ecosystem stability (Alexander et al. 2008; Loewald and Ryan 2020; Manning et al. 2020). Excessive nutrients such as nitrogen and phosphorus can increase the carrying capacity of primary producers in aquatic ecosystems, and this initial spike in algal growth will then increase the abundance of consumers, leading to amplified consumer-resource oscillation cycles, which we term variance-driven destabilization (McCann et al. 2021). This irony that the initial increase in primary productivity could disrupt normal consumer-resource interactions and ultimately destabilize an ecosystem is named the paradox of enrichment (POE; Rosenzweig 1971). In its most extreme form, violent oscillations could drive either consumers or their resources stochastically to extinction.
A number of experimental studies have supported the principle of POE (Rosenzweig 1971; Luckinbill 1973; Veilleux 1979; McCauley et al. 1999; Fussmann et al. 2000; Meyer et al. 2012; Fryxell and Betini 2023; Tadiri et al. 2024), but the complexity in natural food webs can often dampen the destabilizing effect of enrichment through a variety of mechanisms, including predator interference, spatial refugia, and the presence of inedible or unpalatable prey (Otto et al. 2007; Roy and Chattopadhyay 2007; Rall et al. 2008; Feng and Li 2015). In the case where consumers exhibit a varying degree of resource selectivity (Tilman 1982; McCauley and Murdoch 1990; Fryxell and Lundberg 1994; Murdoch et al. 1998; Genkai-Kato and Yamamura 1999), elevated nutrient loading could shift competitive outcomes to favor cyanobacteria and other forms of less edible algae species, at the expense of highly edible forms of green algae (Brooks et al. 2016; Gobler et al. 2017; Trainer et al. 2020), which we term mean-driven destabilization (McCann et al. 2021). In its most extreme form, such mean-driven destabilization can present as competitive displacement, leading to the extinction of green algal species and creating incidence of harmful algal blooms. In Ontario (Canada), the number of confirmed cyanobacteria-dominated algal blooms has increased from less than 5 a year in 1999 to over 60 in 2019, 20 of which were recorded for the first time from new locations (Favot et al. 2023).
While both Rosenzweig’s paradox of enrichment and Tilman’s competitive displacement theory offered meaningful predictions of how nutrient-driven ecosystem instability may arise from local processes, in most freshwater ecosystems; however, instability will also depend on the degree of coupling among local populations occurring across the watershed (Delin and Landon 2002; Loreau et al. 2003; Gounand et al. 2014; Gravel et al. 2016; Laan and Fox 2020; Ryser et al. 2021; Green et al. 2023; Tadiri et al. 2024). This is a critical extension, allowing one to evaluate the impact of enrichment-driven instability as nutrients accumulate across the landscape due to anthropogenic land modification and extreme precipitation events (McCann et al. 2021; Smithwick 2021; Tadiri et al. 2024). Recent spatial nutrient transport theory integrating nutrient-stability theory with meta-ecosystem models (McCann et al. 2021) predicts that downstream accumulation of nutrients and detritus due to fertilizer run-off can amplify both mean- and variance-driven destabilization at a great distance depending on consumers’ feeding preference. When the consumer species consume all resource species, accelerated oscillations in the abundance of resources and consumers will dominate. When the consumer species exhibit selective feeding behaviour and leave inedible species unconsumed, competitive exclusion will drive edible species to functional extinction.
While some empirical studies have provided circumstantial evidence consistent with predictions generated by spatial nutrient transport theory (Delin and Landon 2002; Diaz and Rosenberg 2008; Murphy et al. 2019), controlled experimental studies are uncommon (Laan and Fox 2020; Tadiri et al. 2024). Here, we use time series data collected from an experimental Daphnia-phytoplankton-nutrient model system to test for how different forms of enrichment-driven instability (paradox of enrichment and/or competitive displacement) in a simple three-node network.
Materials and methods
Experimental design
This mesocosm experiment took place in the Hagen Aqualab facility at the University Guelph with 18, 140 L cylindrical fiberglass tanks in a temperature- and light-controlled room (details of this mesocosm setup can be found in Fryxell and Betini 2023) from 17 June to 26 August 2022. Each tank was equipped with an independent LED light system on 10 h:14 h dark: light cycle following a seasonal temperature regime consistent with the growing season in southern Ontario, starting from 18 °C to 24 °C to 18 °C at rate of ± 1.5 °C/week (Fig. S2). We used triple-filtered (25, 1, and 0.2 µm Pentek-pleated filters) and UV-sterilized well water to fill all tanks 17 June 2022, and waited 3 days for the water temperature to reach the initial room temperature of 18 °C. We added 1 g of sodium nitrate (NaNO3), 1.5 mL of liquid fertilizer (Scotts Miracle-Gro LiquaFeed © all-purpose 12-4-8 fertilizer), and 250 million cells of green algae Chlorella vulgaris (strain #90, Canadian Phycological Culture Center) to kickstart the eutrophication experiment, after which we reduced nutrient enrichment to 0.75 mL liquid fertilizer per week per tank, and inoculated 1 g sodium nitrate and 250 million green algae C. vulgaris cells every 2 weeks to all 18 experimental tanks (Fig. S2). The cyanobacteria Microcystis aeruginosa had been used in previous experiments in the tanks and invaded each of the cultures without additional assistance from us. We added ∼330 water fleas (D. magna) to each tank (∼180 juveniles and 150 adults) one week after the initial green algal inoculation.
To simulate a connected landscape, we set up six replicates of a three-node unidirectional network composed of one initial node, one intermediate hub node, and one terminal node. The first serial transfer started 2 weeks after the D. magna inoculation to ensure D. magna populations had successfully colonized each tank. Twice a week thereafter, we disposed of 10% of the liquid (∼14 L) from the terminal tank and then, transferring 10% of the liquid from the initial to the intermediate hub node, and 10% of liquid from the intermediate hub to the terminal node (Fig. 1). We stirred to homogenize contents of all tanks before each transfer to ensure that we captured a representative sample of nutrients, algae, D. magna, and detrital material during each step. After completing transfers, we topped up all tanks with filtered well water to ensure all tanks maintained 140 L capacity at the end of every sampling event. To minimize contamination, each set of replicates had its own polypropylene measuring jug (Catalog #: 470328-332, Avantor, Inc.) and all shared equipment was thoroughly rinsed using filtered water between each replicate. Given that the serial transfers began at the end of week three, we only analyzed data collected after week four, resulting in 252 time series replicates (Fig. S2).
Fig. 1.
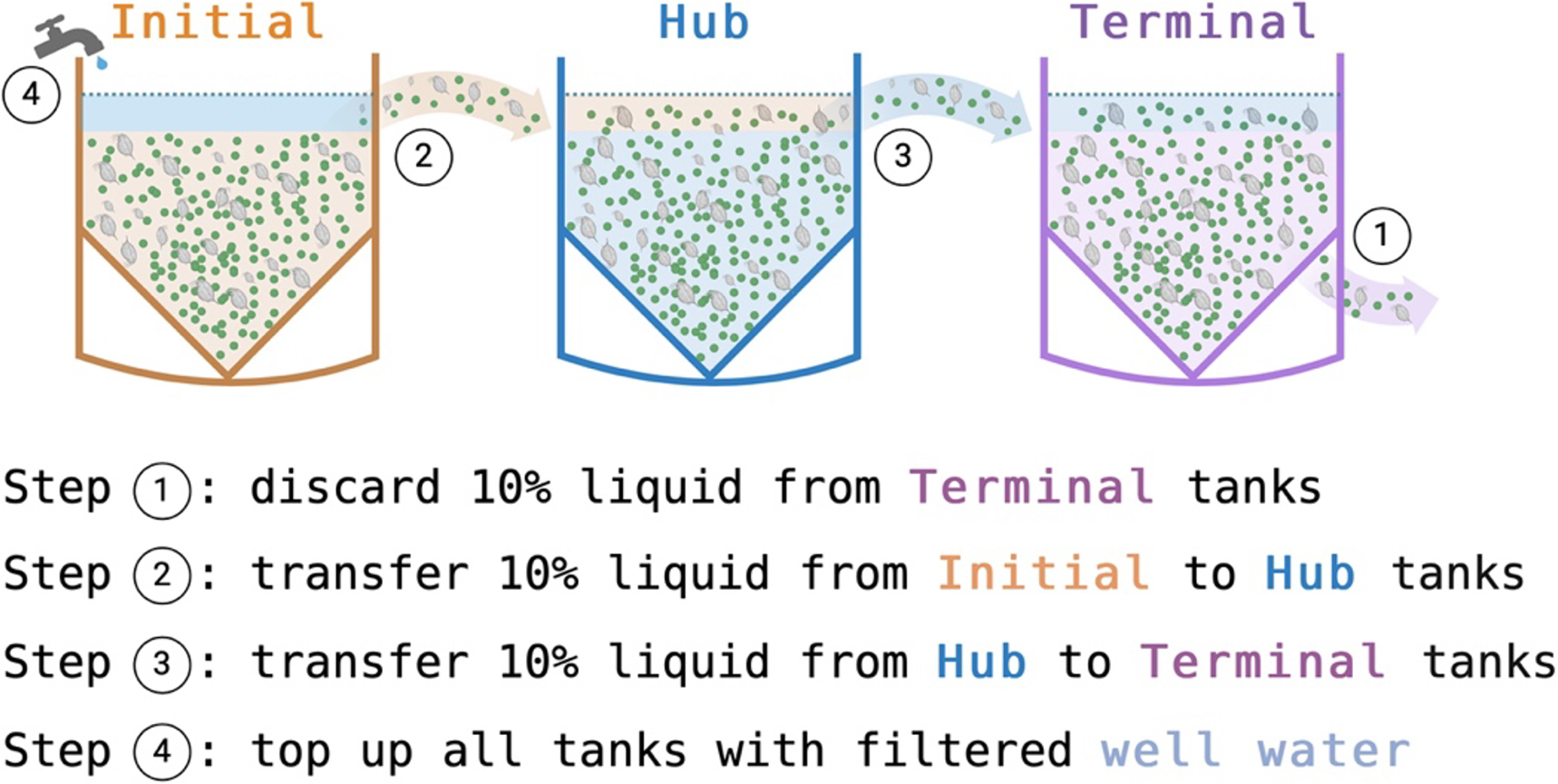
Sample collection and processing
To monitor the changes in nutrient, algal and D. magna density, we sampled each tank twice a week over 10 weeks (20 sampling events per tank, on Tuesdays and Fridays). We collected a 200 mL water sample from each tank and used a shrimp net to systematically sweep D. magna from a total volume of 5.35 L of water. We recorded water temperature and dissolved oxygen (DO) onsite using a hand-held Handy Polaris Probe (OxyGuard Internationals A/S, Denmark), and quantified algal and cyanobacterial density, D. magna density, D. magna biomass, nitrate (N), and total phosphorus concentration (P) on each sampling date.
To estimate algal density, we poured 25 mL of tank water into a glass cuvette and used the benchtop bbe PhycoProbe (bbe Moldaenke GmbH, Germany) with the bbe+++ computer software to record green and cyanobacterial density in cells mL−1. We filtered 30 mL of water through a 0.45 µm Membrane Filter (Millipore Corporation, U.S.A) to remove algae and detrital material and frozen these water chemistry samples until ready for the N and total P analysis. We calorimetrically quantified nitrate using the vanadium reduction method with the Epoch microplate reader (BioTek, VT, USA, Doane and Horwáth 2003), and followed the microplate adaptation of the molybdenum blue method with persulfate digestion to estimate total phosphorus (Murphy and Riley 1962; U.S. Environmental Protection Agency 1993; Ringuet et al. 2011). All water chemistry analyses were run in quadruplicate and results below the detection limit (0.05 mg N L−1 and 0.005 mg P L−1) were considered 0 mg L−1.
To automate the process of obtaining D. magna density and biomass, we used the “Trackdem” R package (Bruijning et al. 2018; Shaw-McDonald 2019). To obtain videos required to use this package, we first carefully poured D. magna samples into separate Petri-dishes. Then, one at a time, each Petri-dish was placed on a lightbox, which was then covered with a custom-made cardboard box with a small window where we secured a smartphone (iPhone X) to take a 15–20 s video (Shaw-McDonald 2019). This set up allowed us to take a video from a fixed distance while minimizing the amount of vibration during filming which would have otherwise interfered with particle tracking in the “Trackdem” algorithm. For samples with high D. magna density (>200 individuals/sample), we divided the sample and took multiple individually labelled videos to reduce the amount of overlapping movement. We then trimmed each video into a short 7 s segment and transformed it into 35 picture frames at 5 frames per second. These pictures were later stacked to create a still background, which allowed the “Trackdem” software to track spatial displacements of particles, their abundance, and biomass in pixels (Bruijning et al. 2018; Shaw-McDonald 2019). Since “Trackdem” assumes each particle to be a perfect circle, we calculated individual D. magna body length L (mm) from the automated output A (pixel) using a calibration curve (), which was then translated into individual body mass (μg) using the formula mass = 4.341*L2.829 (Watkins et al. 2011; Betini et al. 2020). The number and body mass for each D. magna were then added together to create population density (count L−1) and biomass (μg L−1). All D. magna were returned to their tank after all the videos are taken by the end of a sampling day.
Statistical analysis and model
To account for the replication effect and the temporal variation in the time series data, we used mixed-effect models with random intercepts for Date (n = 14) and Replicate (n = 6), combined with pairwise post-hoc tests to determine the effect and the magnitude of nutrient transport on the observed densities of nutrients, detritus, green algae, cyanobacteria, and D. magna across different nodes using the following model structure:where Node is a three-level predictor variable (initial, hub, and terminal), and y represents the suite of response variables including total phosphorus detection probability as a binomial factor (where 0 and 1 indicated a total P concentration below or above detection limit of 0.005 mg.L−1, respectively), total P for samples above detection limit (mg L−1), N (mg L−1), DO (mg L−1), green algal density (cells L−1), cyanobacterial density (cells L−1), D. magna density (counts L−1), and D. magna biomass (μg L−1). We used a mixed-effect generalized logistic regression for total phosphorus detection probabilities and calculated the mean probability () from the estimated odds ratios z (). We specified a fixed reference level of 0 to compare changes in the mean values and provided model estimates with confidence intervals for each node position. We reported significance among initial, hub and terminal node using a pairwise post-hoc test with Holm’s correction (Kuznetsova et al. 2017; Table 1 & Table 2).
(1)
Table 1.
| Total P detection | P concentration | N concentration | DO concentration | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fixed effect | Odds ratios | CI | P | Estimates | CI | P | Estimates | CI | P | Estimates | CI | P |
| Initial node | 0.08 | 0.01–0.60 | – | 0.02 | 0.003 – 0.03 | – | 0.98 | 0.74 – 1.22 | – | 10.82 | 8.42 – 13.22 | – |
| Hub node | 0.39 | 0.06–2.68 | – | 0.03 | 0.02 – 0.04 | – | 1.34 | 1.09 – 1.58 | – | 8.69 | 6.29 – 11.08 | – |
| Terminal node | 1.03 | 0.15–6.96 | – | 0.05 | 0.03 – 0.06 | – | 1.50 | 1.26 – 1.74 | – | 7.80 | 5.40 – 10.20 | – |
| Post-hoc comparison | log-odds | CI | P | Estimates | CI | P | Estimates | CI | P | Estimates | CI | P |
| Hub—initial | 1.57 | 0.28 – 2.87 | 0.009 | 0.01 | 0 – 0.02 | 0.037 | 0.36 | 0.23 – 0.49 | < 0.001 | −2.14 | −2.93 – −1.34 | < 0.001 |
| Terminal—initial | 2.55 | 1.18 – 3.92 | < 0.001 | 0.03 | 0.02 – 0.04 | < 0.001 | 0.52 | 0.39 – 0.65 | < 0.001 | −3.02 | −3.81 – -2.23 | < 0.001 |
| Terminal—hub | 0.98 | −0.15 – 2.1 | 0.042 | 0.02 | 0.01 – 0.03 | < 0.001 | 0.16 | 0.03 – 0.29 | 0.004 | −0.88 | −1.68 – -0.09 | 0.009 |
| Random effects | ||||||||||||
| Residual variance: σ2 | 3.29 | 3.44 × 10−4 | 0.13 | 4.82 | ||||||||
| Random effect variance: τ00 | 9.47 Sample | 2.65 × 10−4Sample | 0.16 Sample | 19.95 Sample | ||||||||
| 0.71 Treatment | 0.95 × 10−4Treatment | 0.01 Treatment | ||||||||||
| Interclass correlation coefficient (ICC) | 0.76 | 0.51 | 0.57 | 0.81 | ||||||||
| 6 Treatment | 6 Treatment | 6 Treatment | ||||||||||
| N | 14 Sample | 12 Sample | 14 Sample | 14 Sample | ||||||||
| Observations | 252 | 93 | 252 | 252 | ||||||||
| Marginal R2/conditional R2 | 0.076/0.774 | 0.154/0.586 | 0.136/0.625 | 0.061/0.817 | ||||||||
Note: Statistical significance (P < 0.05) are highlighted in bold.
Table 2.
| Green algal density | Cyanobacterial density | D. magna density | D. magna biomass | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Predictors | Estimates | CI | P | Estimates | CI | P | Estimates | CI | P | Estimates | CI | P |
| Initial | 16.39 | 16.06–16.73 | – | 15.48 | 15.19 – 15.77 | – | 3.28 | 2.82 – 3.74 | – | 6.20 | 5.82–6.59 | – |
| Hub | 16.60 | 16.27–16.94 | – | 15.58 | 15.29 – 15.87 | – | 3.62 | 3.16 – 4.08 | – | 6.48 | 6.10 – 6.87 | – |
| Terminal | 16.67 | 16.34–17.0 | – | 15.51 | 15.22 – 15.80 | – | 3.56 | 3.10 – 4.02 | – | 6.39 | 6.00 – 6.77 | – |
| Post-hoc comparison | Estimates | CI | P | Estimates | CI | P | Estimates | CI | P | Estimates | CI | P |
| Hub—initial | 0.21 | 0.08 – 0.33 | < 0.001 | 0.1 | −0.06 – 0.26 | 0.46 | 0.33 | 0.15 – 0.53 | < 0.001 | 0.28 | 0.07 – 0.48 | 0.006 |
| Terminal—initial | 0.28 | 0.15 – 0.4 | < 0.001 | 0.03 | −0.13 – 0.20 | 0.67 | 0.28 | 0.09 – 0.47 | < 0.001 | 0.18 | −0.03 – 0.39 | 0.08 |
| Terminal—hub | 0.07 | −0.06 – 0.2 | 0.202 | −0.07 | −0.23 – 0.09 | 0.64 | −0.06 | −0.24 – 0.13 | 0.47 | −0.10 | −0.31 – 0.12 | 0.29 |
| Random effects | ||||||||||||
| σ2 | 0.13 | 0.21 | 0.26 | 0.34 | ||||||||
| τ00 | 0.34 Sample | 0.19 Sample | 0.72 Sample | 0.47 Sample | ||||||||
| 0.02 Treatment | 0.03 Treatment | 0.001 Treatment | 0.006 Treatment | |||||||||
| ICC | 0.74 | 0.52 | 0.73 | 0.58 | ||||||||
| N | 6 Treatment | 6 Treatment | 6 Treatment | 6 Treatment | ||||||||
| 14 Sample | 14 Sample | 14 Sample | 14 Sample | |||||||||
| Observations | 252 | 252 | 252 | 252 | ||||||||
| marginal R2 /conditional R2 | 0.028/0.746 | 0.004/0.518 | 0.022/0.737 | 0.016/0.591 | ||||||||
Note: Statistical significance (P < 0.05) are highlighted in bold.
In addition to comparing the observed means across the network, we calculated per capita per day rates of exponential growth (, where τ is the time between consecutive sampling events, either three or four days) for green algae (r 1), cyanobacteria (r 2), and D. magna (r 3) to evaluate the effect of serial transfer location (three-level factor: initial, hub and terminal), abiotic (P (mg.L−1), N (mg.L−1), DO (mg.L−1), temperature (°C)), and biotic factors (green algae (N1 : cells.L−1), cyanobacteria (N2 : cells.L−1), D. magna density (N3 : counts.L−1), and D. magna biomass) on the rates of population change in a mix-effect linear model. Since D. magna density and D. magna biomass were highly correlated, we only used the residuals from (, F1,232 = 910.4, R2 = 0.8, P < 0.001) to represent D. magna biomass. We checked the collinearity of all predictor variables using variance inflation factor (GVIF1/(2*DF)) and all variables were below the collinearity threshold of 2.5 (Johnston et al. 2018). Since this set of full models incorporated all variables (Table S1), we applied a backward elimination method to reduce non-significant effects (Chambers and Hastie 2017; Kuznetsova et al. 2017). Key factors that influenced the rate of change for green algae (r 1), cyanobacteria (r 2), and D. magna (r 3) were as follows:
(2)
where ai and bi represent the density-dependent intra- and inter-specific competition (ai, bi < 0), the intercepts c i represent maximum per capita growth rate of species i, and d represents the positive correlation between green algae and D. magna growth rate.
To further evaluate the competitive outcome between green algae and cyanobacteria occurred in our system, we parameterized the following Lotka–Volterra competition model (Lotka 1925; Volterra 1926):where N1 and N2 represent green algal and cyanobacterial density, rmax1 and rmax2 depict their maximum growth rates (rmaxi = ci), K1 and K2 represent their carrying capacities (Ki = − ci/ai). The Lotka–Volterra competition coefficient α12 indicates the per capita impact of cyanobacteria on the population growth of green algae (α12 = b1/a1), and α21 describes the impact of green algae on the population growth of cyanobacteria (α21 = b2/a2). We used the average green algal and cyanobacterial density from sample #7 (N1, 0 = 5.67 × 107 ; N2,0 = 2.75 × 106 cell L−1) as the initial green algal and cyanobacterial density and simulated their population trajectory for 120 days and calculated the equilibrium density (where , ). The competitive outcome between green algae and cyanobacteria depends on both their respective carrying capacity and their competition coefficients, expressed as follows:
(3)
, : two species coexist but the equilibrium is unstable;
, : green algae out-competes cyanobacteria;
, : cyanobacteria out-competes green algae;
, : two species coexist and the equilibrium is stable.
All statistical analyses were conducted in R 2023.09.1 + 494 (R Core Team 2023). The linear mixed-effect models were fitted by maximum likelihood using the package “lme4” (Bates et al. 2015) and model coefficients were visualized using package “sjPlot” (Lüdecke 2023). Post-hoc analysis was done using the package “multcomp” and P-values were reported with Holm’s correction (Hothorn et al. 2008). Model optimization was performed using “lmerTest” package (Kuznetsova et al. 2017). All figures were created using the package “ggplot2” (Allen et al. 2019) with colour palette from “RColorBrewer” (Neuwirth 2022) and “ggtext” (Wilke and Wiernik 2022). The variance inflation factor was calculated using the package “car” (Fox and Weisberg 2019). All data and R scripts used for the analyses is made publicly available on GitHub public repository in the Data Availability Statement.
Results
We compared the observed mean densities of nutrients, detritus load, green algae, cyanobacteria, and D. magna across the initial, hub, and terminal nodes. Nutrients showed significant evidence of downstream accumulation consistent with predictions from spatial nutrient transport theory: terminal nodes had the highest detectability and concentration of total P (p̂ = 0.51, CI = 0.13–0.87, Fig. 2a; β = 0.05, CI = 0.03–0.06, Fig. 2b; Table 1) and N on average (β = 1.50, CI = 1.26–1.74, Fig. 2c, Table 1), significantly higher than those of hub and initial nodes (P < 0.05, Table 1). We found that node position explained 15.4% and 13.6% of the variation in the observed mean total P and total N concentration (Table 1), while random effect of time and replication contributed to an additional 43% and 49% of the variation (conditional-marginal R2; Figs. 2d and 2e). This highlighted that spatiotemporal factors could contribute to as much as 62% of the observed variation in nutrient concentrations.
Fig. 2.
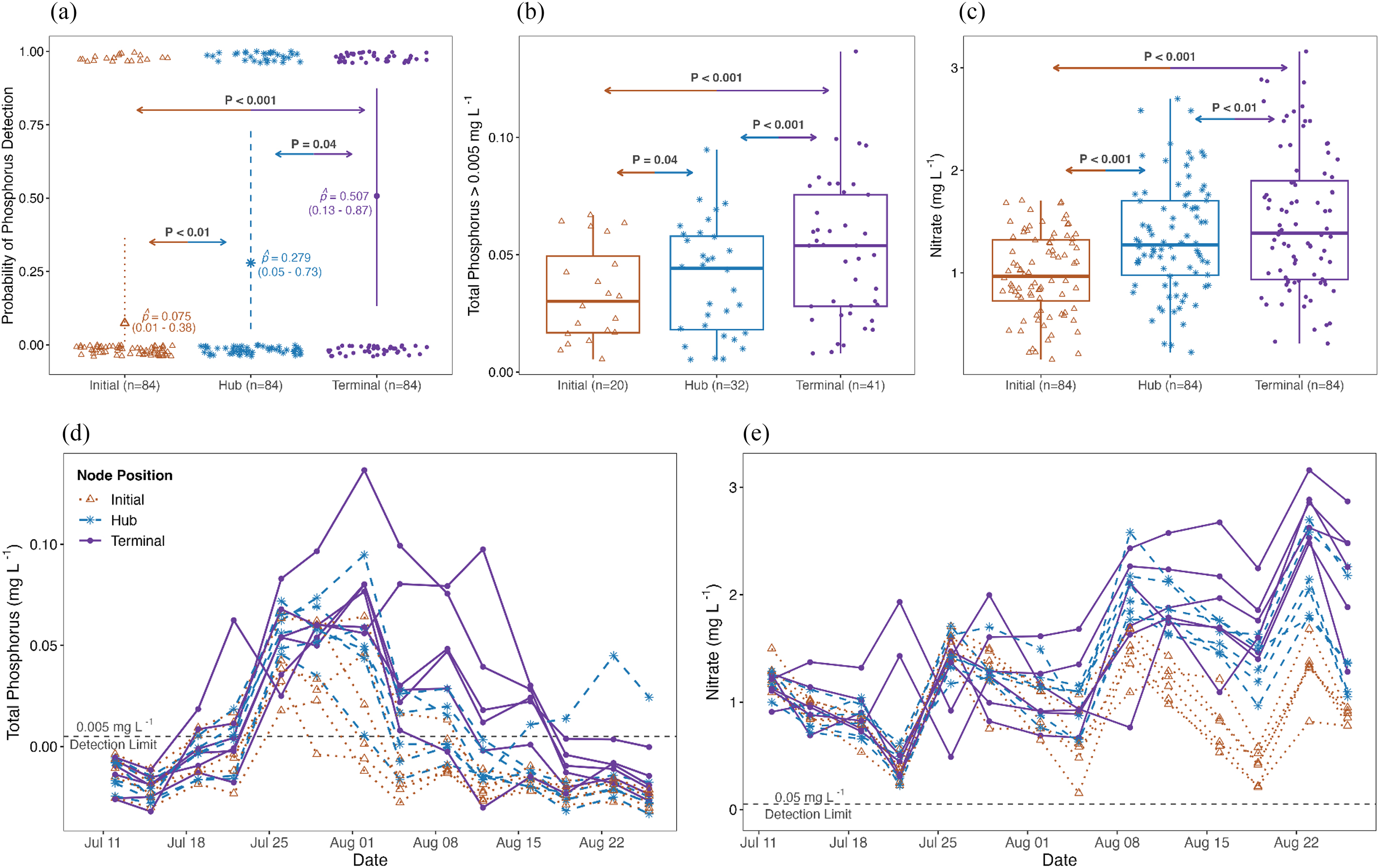
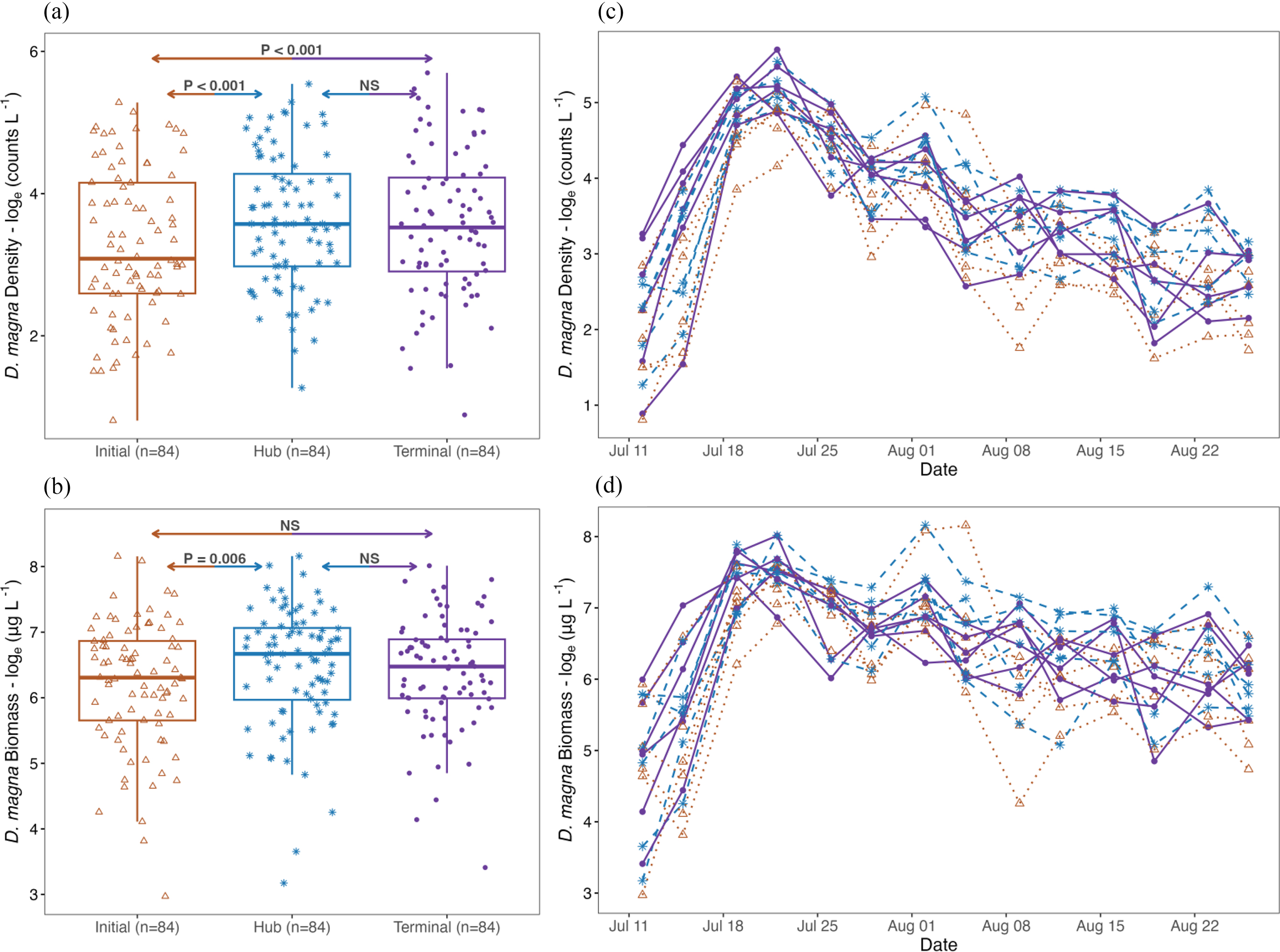
Increased levels of nutrients were associated with modest increases in the abundance of green algae and D. magna in hub and terminal nodes relative to the initial node (Figs. 3 and 4). While we found no difference between hub and terminal nodes (P > 0.05, Table 2), the initial node had the lowest densities of green algae (β = 15.48, CI = 15.19–15.77, Table 2, Fig. 3a), D. magna (β = 3.26, CI = 2.78–3.73, Table 2, Fig. 3a), and D. magna biomass (β = 7.48, CI = 7.09–7.88, Table 2, Fig. 3b). There was little evidence that cyanobacteria density responded to serial transfer, with no significant difference detected among initial, hub, or terminal nodes (P > 0.05, Table 2, Fig. 3b).
Fig. 3.

Fig. 4.
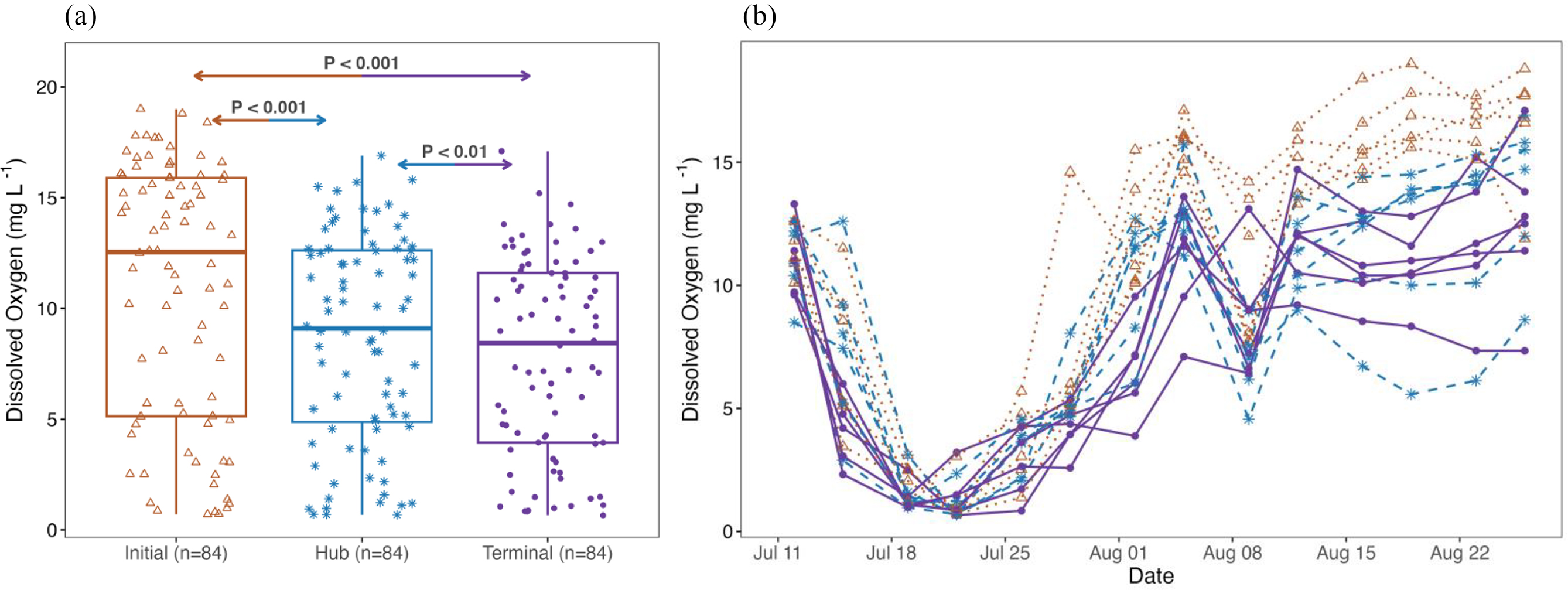
While we observed modest increase in algal and D. magna density along the spatial gradient, we found strong evidence of reduced DO concentration downstream and recorded the lowest DO concentration in the terminal nodes (β = 7.80, CI = 5.40–10.20, Table 1, Fig. 5). Given that green algae and cyanobacterial cell density, D. magna abundance, and detrital materials are all known to affect oxygen production and uptake, changes in DO concentration could be due to multiple causes. In our system where phytoplankton or zooplankton showed no difference between the hub and terminal nodes (P > 0.05, Table 2), we suggest that the decreasing DO was likely driven by increased detritus load and bacteria-driven oxygen uptake.
Fig. 5.
Tank position did not influence the exponential growth rates recorded for green algae or cyanobacteria (P > 0.05, Table S1), but exponential growth rates within tanks consistently reflected localized effects of intra- and inter-specific competition (Table 3). Green algal growth rate showed negative density dependence (a1 = −3.00 × 10−9, CI = (−3.52 – −2.47) × 10−9, P < 0.001, Fig. 6a), and strong negative inter-specific competition with cyanobacteria (b1 = −5.73 x10−9, CI = (−9.59 – −1.88) × 10−9, P = 0.004, Fig. 6c). Similarly, cyanobacterial growth rate decreased with cyanobacteria density (a2 = −1.82 × 10−9, CI = (−2.48 to −1.15) × 10−9, P < 0.001, Fig. 6b) and green algae density (b2 = −19.32 × 10−9, CI = (−24.59 to −14.1) × 10−9, P < 0.001, Fig. 6d). D. magna growth rate demonstrated strong density dependence (a3 = −0.0023, CI = −0.003 to −0.002, P < 0.001, Table 3) and a positive response to increasing green algae abundance (b3 = 1.15 × 10−9, CI = (0.34–1.95) × 10−9, P = 0.005, Table 3), but no apparent relationship to cyanobacteria abundance. Abiotic factors such as total P, N, DO, and temperature had no detectable influence on exponential growth rates (P > 0.05, Table S1).
Fig. 6.
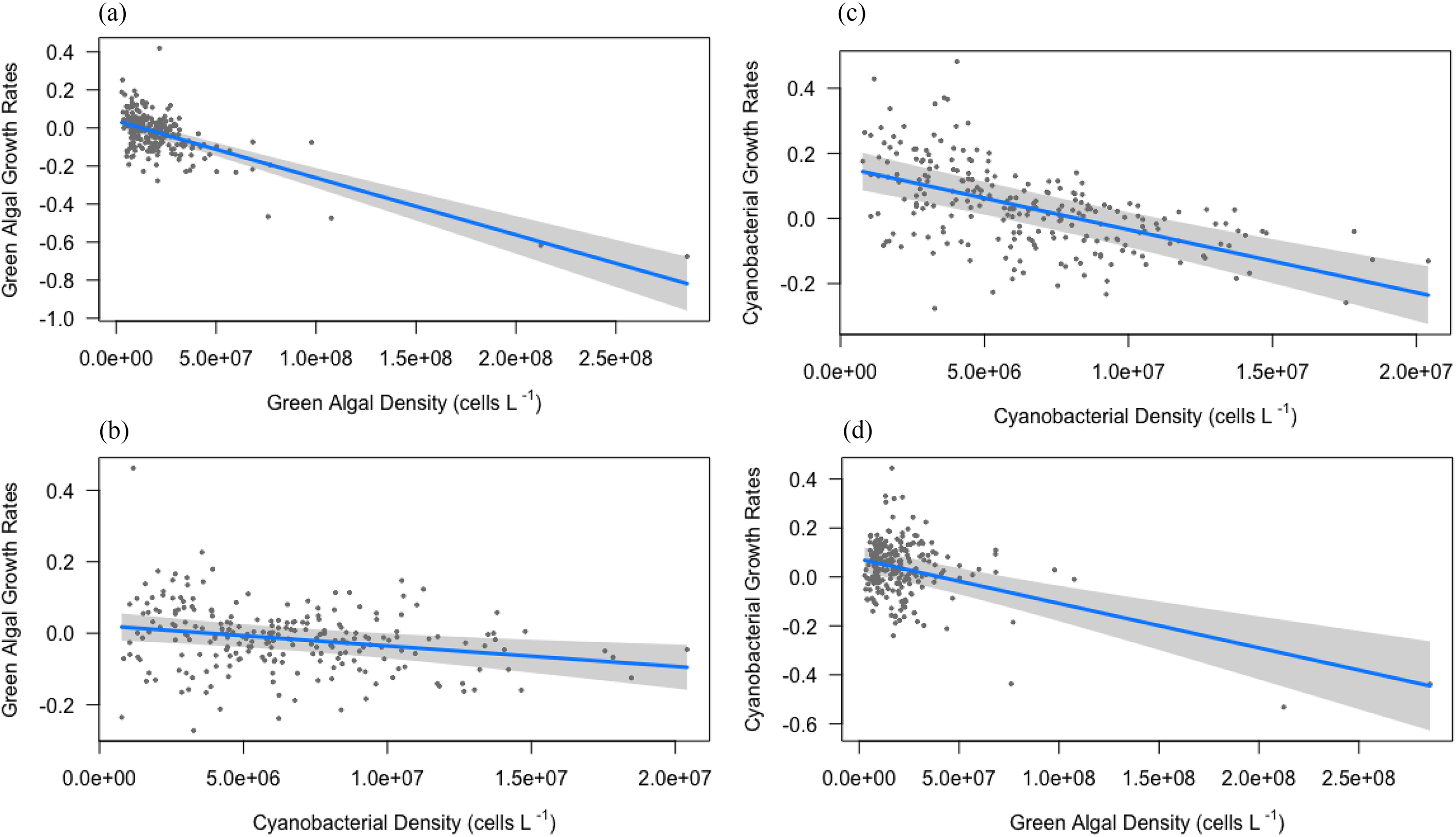
Table 3.
| Green algal growth rates | Cyanobacterial growth rates | D. magna growth rates | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Predictors | Estimates | CI | p | Estimates | CI | p | Estimates | CI | P |
| rmax | 0.071 | 0.03 – 0.11 | 0.001 | 0.19 | 0.13 – 0.25 | < 0.001 | 0.106 | 0.01 – 0.21 | 0.051 |
| Green algal density (cells L−1) | −3.00 (×10−9) | −3.52 to −2.47 (×10−9) | < 0.001 | −1.82 (×10−9) | −2.48 to −1.15 (×10−9) | < 0.001 | 1.15 (×10−9) | 0.34 – 1.95 (×10−9) | 0.005 |
| Cyanobacterial density (cells L−1) | −5.73 (×10−9) | −9.59 to −1.88 (×10−9) | 0.004 | −19.32 (×10−9) | −24.5 to −14.1 (×10−9) | < 0.001 | |||
| D. magna density (counts L−1) | −0.0023 | −0.003 to –0.002 | < 0.001 | ||||||
| Random effects | |||||||||
| σ2 | 0.008 | 0.012 | 0.018 | ||||||
| τ00 | 0.003 Sample | 0.005 Sample | 0.027 Sample | ||||||
| 0.001 Treatment | |||||||||
| ICC | 0.27 | 0.33 | 0.60 | ||||||
| N | 6 Treatment | ||||||||
| 13 Sample | 13 Sample | 13 Sample | |||||||
| Observations | 234 | 234 | 234 | ||||||
| Marginal R2 /conditional R2 | 0.373/0.546 | 0.272/0.513 | 0.247/0.696 | ||||||
Note: Regression estimates from full models can be found in Table S1. Statistical significance (P < 0.05) are highlighted in bold.
Coefficients of the Lotka–Volterra competition model suggest that even though cyanobacteria exhibited higher competitive inhibition on green algae than the converse (α12 = 1.87 vs. α21 = 0.094, Table 4, Fig. 7), stability analysis indicated a locally stable equilibrium with trajectories for both green algae and cyanobacteria converging on the interior intersection of their respective null isoclines (Fig. 7b).
Fig. 7.
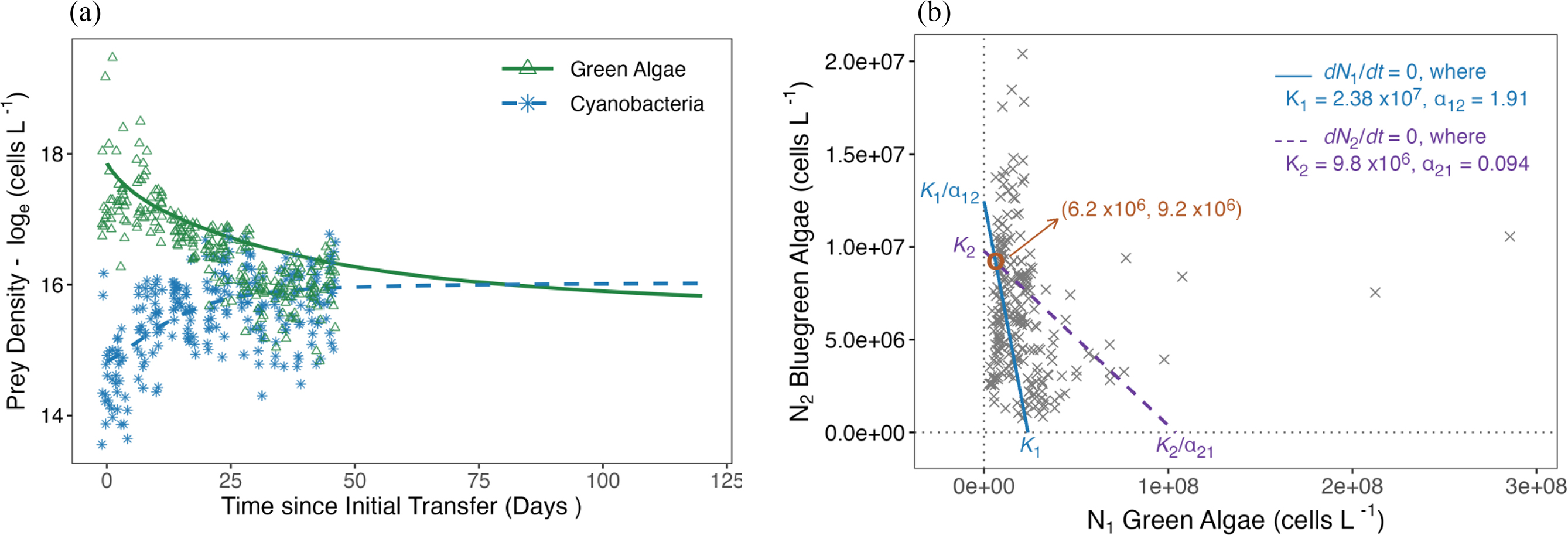
Table 4.
| Lotka–Volterra competition parameters | |||
|---|---|---|---|
| Parameter | Descriptions | Value | |
| Green algae | rmax1 | Maximum growth rate | 0.071 |
| α21 | Inhibition from cyanobacteria | 1.91 | |
| K1 | Carrying capacity (cells L−1) | 2.38 × 107 | |
| Equilibrium density (cells L−1) | 6.2 × 106 | ||
| Cyanobacteria | rt2 | Maximum growth rate | 0.19 |
| α12 | Inhibition from green algae | 0.094 | |
| K2 | Carrying capacity (cells L−1) | 9.8 × 106 | |
| Equilibrium density (cells L−1) | 9.2 × 106 | ||
| Stability analysis | |||
| K2 = 9.8 × 106 < K1/α12 = 1.24 × 107 | |||
| K1 = 2.38 × 107 < K2/α21 = 1.04 × 108 | |||
Discussion
From heavy usage of synthetic fertilizers and pesticide to large scale land clearing and modification, intense agricultural production is increasingly recognized as a key threat to aquatic ecosystem services (Tilman 1999; Dudley and Alexander 2017). Over half of nitrogen and phosphorus inputs from fertilizer application to farmlands can be lost to surrounding terrestrial and aquatic ecosystems, destabilizing food web structure, interrupting nutrient cycling, and reducing ecosystem functions (Bouwman et al. 2009; Zou et al. 2022). In interconnected watersheds, nutrients can travel great distances, affecting many coupled ecosystems far from the source (Loreau et al. 2003; Gounand et al. 2014; McCann et al. 2021; Tadiri et al. 2024). To test the impact of nutrient transport on the presence and magnitude of enrichment-driven instabilities across a spatial gradient, we used the classic Daphnia-phytoplankton-nutrient model system in a three-node unidirectional mesocosm experiment.
We found that even though nitrate, total phosphorus, and detrital materials accumulated significantly downstream (Figs. 2 and 5), a pattern consistent with predictions from the spatial nutrient transport theory (McCann et al. 2021), this did not result in extreme amplification of population abundance, cycling, or complete competitive exclusion of green algae in hub or terminal nodes (Figs. 3c–3d and 4c–4d). Instead, our algal/cyanobacteria community converged on a stable equilibrium, where green algae and cyanobacteria co-exist at around 6.2, and 9.2 million cells L−1, respectively, independent from node position (Fig. 7b). Here, we discussed several factors at work in our system that could contribute to the stability in our system.
First, the enrichment level in our system could be inadequate. Given the detection limit of 0.005 mg P L−1, we were only able to quantify total phosphorus in 37% of our total samples (n = 92), 40% of which were below the blooming threshold of 0.03 mg L−1 (n = 36, Dodds et al. 2002). Even though we added liquid fertilizer weekly (∼15.8 mg P and 50 mg N Tank−1), we found phosphorus uptake to be extremely rapid and that the phosphorus level sometimes dropped below the detection limit within 24 hours. Compared to the ideal 16:1 N:P ratio for algal growth, the exceedingly high stoichiometric ratio of N:P in our system (48:1 ± 43:1, n = 92) suggested that phosphorus was the limiting nutrient in our system (Guildford and Hecky 2000), which could lower algal growth rate (Li et al. 2022) and reduce the potential for complete competitive exclusion (Redfield 1934; Takamura et al. 1992; Neill 2005).
Second, strong intra- and inter-specific competition between green algae and cyanobacteria could be stabilizing. Previous work suggests that nutrient enrichment tends to amplify Daphnia-phytoplankton prey-escape cycles most readily in simple systems composed of strictly edible algal species (McCauley et al. 1999). In natural studies or experiments where less edible or unpalatable prey are present, enrichment rarely leads to prey-escape cycles (McCauley and Murdoch 1990; Murdoch et al. 1998; Genkai-Kato and Yamamura 1999; McCauley et al. 1999; Mougi and Nishimura 2007), as a result of interspecific competition among algal prey (Kretzschmar et al. 1993; Genkai-Kato and Yamamura 1999) and/or changes in predator foraging (McCauley et al. 1988, 1999; Mougi and Nishimura 2007; Yin et al. 2010). Similarly, we found that intra- and inter-specific competition between green algae and cyanobacteria were key factors influencing algal growth rates, but not D. magna abundance (P > 0.05, Table S1). Cyanobacteria had higher maximum rates of increase (r2 = 0.19 > r1 = 0.071, Table 4), suggesting they were more effective at nutrient uptake than green algae. Cyanobacteria exerted stronger evidence of competitive inhibition on the growth of green algae than vice versa (α21 = 1.87 > α12 = 0.094, Table 4). While competitive displacement by cyanobacteria did not drive green algae to complete extinction (McCann et al. 2021), it depressed long-term levels of green algal abundance to a considerable extent. Cyanobacteria reached a 50% higher density at equilibrium than green algae (9.2 × 106 cells L−1 vs. 6.2 × 106 cells L−1). This is particularly impressive in relation to the substantial difference in their respective carrying capacities (9.8 × 106 cells L−1 vs. 2.38 × 107 cells L−1).
Third, all our tanks were subject to augmented inoculations of green algae every two weeks, which could also contribute to the persistence of green algae in our system. We note that cyanobacteria slowly, and successfully, invaded our mesocosms 3 weeks after green algae density had already established population densities of 2.1 ± 1.2 × 105 cells L−1. It is conceivable that algal competitive outcomes might change if initial levels of abundance or temperature conditions had been different. Slow rates of cyanobacteria invasion might be expected, because per capita growth rates of M, aeruginosa are much lower at 18 °C than at 25 °C (Shaw 2020), unlike green algae C. vulgaris (Jarvis et al. 2016).
Lastly, difference in prey palatability could influence D. magna’s feeding behaviour. Our results showed that D. magna population growth rates were positively related only to green algae abundance, and there was no detectable influence of cyanobacteria abundance. This suggested that while cyanobacteria in our system are edible, they were so poorly profitable that they failed to fulfill D. magna’s energetic needs. Bench-top trials conducted in our lab were consistent with this hypothesis, showing that per capita rates of survival, reproduction, and individual mass gain by D. magna declined with an increasing ratio of M, aeruginosa to C. vulgaris in their diet (Shahmohamadloo et al. 2022). Interestingly, the lack of oscillatory predator-prey cycle from our experiment were consistent with predictions presented by Genkai-Kato and Yamamura using foraging theory (1999). In their predator-prey model, one predator could consume two prey species of varying profitability (Figs. 1a-4), and when the less palatable prey species approached the critical profitability threshold, oscillations were nearly muted. A similar theoretical prediction of behavioral stabilization comes from Fryxell and Lundberg's (1994) consumer-resource models with optimal foraging by a single consumer species and two resource species, the least profitable of which is unable to sustain predators on its own. Prey escape cycles can also be muted by resource limitation (Carroll et al. 2022; Luckinbill 1973), complexity of natural food webs (Rall et al. 2008; Kretzschmar et al. 1993; McCauley et al. 1999; Mougi and Nishimura 2007; Murdoch et al. 1998; Persson et al. 2001; Roy and Chattopadhyay 2007), landscape connectivity (Feng and Li 2015; Gounand et al. 2014; Gravel et al. 2016; Tadiri et al. 2024), behavioural plasticity (McCauley and Murdoch 1990; Mougi and Nishimura 2008), and rates of transport between interconnected ecosystems (Gounand et al. 2014; McCann et al. 2021).
There are some limitations to our study. It should be noted that our experimental design utilized a linear structure rather than a dendritic configuration (McCann et al. 2021). We recognized that linear network would likely promote instability due to the lack of asynchronous input (Anderson and Hayes 2018; Tadiri et al. 2024), and we are cautious that the enrichment level in our system might not reflect the level of enrichment in natural riverine systems. Our system has two sources of enrichment: downstream accumulation from directional movement of water, nutrient, resource, consumers, and detritus, as well as weekly enrichment of fertilizers. Future work could explore how different enrichment level may interact with landscape configurations (linear vs. dendritic) to influence meta-ecosystem stability. Additionally, sexual production of ephippia in Daphnia could influence consumer-resource dynamics (McCauley et al. 1999), and would be worth further investigation. Finally, our experiment followed a seasonal temperature regime, coupled with a constant flow rate where all nodes had the same initial conditions. In natural environment, coupled ecosystems would exhibit a higher degree of heterogeneity of varying species composition and climatic condition, which could be another important aspect for future work.
Eutrophication from agriculture runoff remains a persistent and complex problem. As nutrients rapidly travel across coupled ecosystems via increasingly interconnected landscape (Gravel et al. 2016; Loreau et al. 2003; McCann et al. 2021; Smithwick 2021), it is essential to understand the effect and magnitude of nutrient transport on food web stability. Consistent with the recent spatial nutrient transport theory (McCann et al. 2021), our Daphnia-phytoplankton-nutrient model system provided evidence of strong nutrient and detritus accumulation downstream. However, these changes did not lead to an amplified Daphnia-phytoplankton paradox cycle or complete competitive replacement of green algae by less edible cyanobacteria. Instead, our system converged on a stable coexisting equilibrium where cyanobacteria inhibited green algal growth and suppressed green algae abundance to a quarter of its normal carrying capacity. Improved stability in our system could result from nutrient limitation, prey competition and palatability, and weekly inoculation of green algae.
Acknowledgements
We thank all Aqualab staff members: Matt Cornish, Carolyn Trombley, and Mike Davies, who supported all technical aspects of this experiment. We are grateful for Matthew Pavusa and Dr. Andrew McDougall for providing training and assistance on our water chemistry analysis.
References
Alexander R.B., Smith R.A., Schwarz G.E., Boyer E.W., Nolan J.V., Brakebill J.W. 2008. Differences in phosphorus and nitrogen delivery to The Gulf of Mexico from the Mississippi River Basin. Environmental Science & Technology, 42, 822–830.
Allen M., Poggiali D., Whitaker K., Marshall T.R., Kievit R.A. 2019. Raincloud plots: a multi-platform tool for robust data visualization. Wellcome Open Research, 4, 63.
Anderson K.E., Hayes S.M. 2018. The effects of dispersal and river spatial structure on asynchrony in consumer–resource metacommunities. Freshwater Biology, 63, 100–113.
Bates D., Mächler M., Bolker B., Walker S. 2015. Fitting linear mixed-effects models using lme4. Journal of Statistical Software, 67.
Betini G.S., Wang X., Avgar T., Guzzo M.M., Fryxell J.M. 2020. Food availability modulates temperature-dependent effects on growth, reproduction, and survival in Daphnia magna. Ecology and Evolution, 10, 756–762.
Bouwman A.F., Beusen A.H.W., Billen G. 2009. Human alteration of the global nitrogen and phosphorus soil balances for the period 1970–2050. Global Biogeochemical Cycles, 23, 2009GB003576.
Brooks B.W., Lazorchak J.M., Howard M.D.A., Johnson M.-V.V., Morton S.L., Perkins D.A.K., et al. 2016. Are harmful algal blooms becoming the greatest inland water quality threat to public health and aquatic ecosystems?: harmful algal blooms: the greatest water quality threat? Environmental Toxicology and Chemistry, 35, 6–13.
Bruijning M., Visser M.D., Hallmann C.A., Jongejans E. 2018. trackdem : automated particle tracking to obtain population counts and size distributions from videos in r. Methods in Ecology and Evolution, 9, 965–973.
Carroll O., Batzer E., Bharath S., Borer E.T., Campana S., Esch E., et al. 2022. Nutrient identity modifies the destabilising effects of eutrophication in grasslands. Ecology Letters, 25, 754–765.
Chambers J.M., Hastie T.J. (Eds.) 2017. Statistical Models in S, 1st ed. Routledge.
Delin G.N., Landon M.K. 2002. Effects of surface run-off on the transport of agricultural chemicals to ground water in a sandplain setting. Science of The Total Environment, 295, 143–155.
Diaz R.J., Rosenberg R. 2008. Spreading dead zones and consequences for marine ecosystems. Science, 321, 926–929.
Doane T.A., Horwáth W.R. 2003. Spectrophotometric determination of nitrate with a single reagent. Analytical Letters, 36, 2713–2722.
Dodds W.K., Smith V.H., Lohman K. 2002. Nitrogen and phosphorus relationships to benthic algal biomass in temperate streams. Canadian Journal of Fisheries and Aquatic Sciences, 59, 865–874.
Dudley N., Alexander S. 2017. Agriculture and biodiversity: a review. Biodiversity, 18, 45–49.
FAO. 2018. The future of food and agriculture—alternative pathways to 2050.
FAO. 2022. FAOSTAT: Cropland nutrient budget. Retrieved December 12, 2023 from https://www.fao.org/faostat/en/#data/ESB.
Favot E.J., Holeton C., DeSellas A.M., Paterson A.M. 2023. Cyanobacterial blooms in Ontario, Canada: continued increase in reports through the 21st century. Lake and Reservoir Management, 39, 1–20.
Feng Z.C., Li Y.C. 2015. A resolution of the paradox of enrichment. International Journal of Bifurcation and Chaos, 25, 1550094.
Fox J., Weisberg S. 2019. An R companion to applied regression. 3rd ed. Sage, Thousand Oaks, CA.
Fryxell J.M., Betini G.S. 2023. Algal blooms as a reactive dynamic response to seasonal perturbation in an experimental system. Theoretical Ecology, 16, 151–160.
Fryxell J.M., Lundberg P. 1994. Diet choice and predator-prey dynamics. Evolutionary Ecology, 8,407–421.
Fussmann G.F., Ellner S.P., Shertzer K.W., Hairston N.G. Jr. 2000. Crossing the hopf bifurcation in a live predator-prey system. Science 290, 1358–1360.
Genkai-Kato M., Yamamura N. 1999. Unpalatable prey resolves the paradox of enrichment. Proceedings of the Royal Society of London. Series B: Biological Sciences, 266, 1215–1219.
Gobler C.J., Doherty O.M., Hattenrath-Lehmann T.K., Griffith A.W., Kang Y., Litaker R.W. 2017. Ocean warming since 1982 has expanded the niche of toxic algal blooms in the North Atlantic and North Pacific oceans. Proceedings of the National Academy of Sciences, 114, 4975–4980.
Gounand I., Mouquet N., Canard E., Guichard F., Hauzy C., Gravel D. 2014. The paradox of enrichment in metaecosystems. The American Naturalist, 184, 752–763.
Gravel D., Massol F., Leibold M.A. 2016. Stability and complexity in model meta-ecosystems. Nature Communications, 7, 12457.
Green M.D., Woodie C.A., Whitesell M., Anderson K.E., 2023. Long transients and dendritic network structure affect spatial predator–prey dynamics in experimental microcosms. Journal of Animal Ecology, 92, 1416–1430.
Guildford S.J., Hecky R.E. 2000. Total nitrogen, total phosphorus, and nutrient limitation in lakes and oceans: is there a common relationship? Limnology & Oceanography, 45, 1213–1223.
Hothorn T., Bretz F., Westfall P. 2008. Simultaneous inference in general parametric models. Biometrical Journal, 50, 346–363.
Jarvis L., McCann K., Tunney T., Gellner G., Fryxell J.M. 2016. Early warning signals detect critical impacts of experimental warming. Ecology and Evolution, 6, 6097–6106.
Johnston R., Jones K., Manley D. 2018. Confounding and collinearity in regression analysis: a cautionary tale and an alternative procedure, illustrated by studies of British voting behaviour. Quality & Quantity, 52, 1957–1976.
Kretzschmar M., Nisbet R.M., Mccauley E. 1993. A predator-prey model for zooplankton grazing on competing algal populations. Theoretical Population Biology, 44, 32–66.
Kuznetsova A., Brockhoff P.B., Christensen R.H.B. 2017. lmerTest package: tests in linear mixed effects models. Journal of Statistical Software, 82.
Laan E., Fox J.W. 2020. An experimental test of the effects of dispersal and the paradox of enrichment on metapopulation persistence. Oikos, 129, 49–58.
Li M., Li Y., Zhang Y., Xu Q., Iqbal M.S., Xi Y., Xiang X. 2022. The significance of phosphorus in algae growth and the subsequent ecological response of consumers. Journal of Freshwater Ecology, 37, 57–69.
Loewald A., Ryan P. 2020. A review of phosphorous and nitrogen in groundwater and lakes(Vermont Geological Survey Technical Report).
Loreau M., Mouquet N., Holt R.D. 2003. Meta-ecosystems: a theoretical framework for a spatial ecosystem ecology. Ecology Letters, 6, 673–679.
Lotka A.J. 1925. Elements of physical biology. Nature, 116, 461–461.
Luckinbill L.S. 1973. Coexistence in laboratory populations of Paramecium Aurelia and its predator didinium Nasutum. Ecology, 54, 1320–1327.
Lüdecke D. 2023. sjPlot: data visualization for statistics in Social science.
Manning D.W.P., Rosemond A.D., Benstead J.P., Bumpers P.M., Kominoski J.S. 2020. Transport of N and P in U.S. streams and rivers differs with land use and between dissolved and particulate forms. Ecological Applications, 30, e02130.
McCann K.S., Cazelles K., MacDougall A.S., Fussmann G.F., Bieg C., Cristescu M., et al. 2021. Landscape modification and nutrient-driven instability at a distance. Ecology Letters, 24, 398–414.
McCauley E., Murdoch W.W. 1990. Predator–prey dynamics in environments rich and poor in nutrients. Nature, 343, 455–457.
McCauley E., Murdoch W.W., Watson S. 1988. Simple models and variation in plankton densities among lakes. The American Naturalist, 132, 383–403.
McCauley E., Nisbet R.M., Murdoch W.W., De Roos A.M., Gurney W.S.C. 1999. Large-amplitude cycles of Daphnia and its algal prey in enriched environments. Nature, 402,653–656.
Meyer K.M., Vos M., Mooij W.M., Hol W.H.G., Termorshuizen A.J., Van Der Putten W.H. 2012. Testing the paradox of enrichment along a land use gradient in a multitrophic aboveground and belowground community. PLoS ONE, 7, e49034.
Mogollón J.M., Beusen A.H.W., Van Grinsven H.J.M., Westhoek H., Bouwman A.F. 2018. Future agricultural phosphorus demand according to the shared socioeconomic pathways. Global Environmental Change, 50, 149–163.
Mougi A., Nishimura K. 2007. A resolution of the paradox of enrichment. Journal of Theoretical Biology, 248, 194–201.
Mougi A., Nishimura K. 2008. The paradox of enrichment in an adaptive world. Proceedings of the Royal Society B: Biological Sciences, 275, 2563–2568.
Murdoch W.W., Nisbet R.M., McCauley E., deRoos A.M., Gurney W.S.C. 1998. Plankton abundance and dynamics across nutrient levels: tests of hypotheses. Ecology, 79, 1339–1356.
Murphy G.E.P., Wong M.C., Lotze H.K. 2019. A human impact metric for coastal ecosystems with application to seagrass beds in Atlantic Canada. FACETS, 4, 210–237.
Murphy J., Riley J.P. 1962. A modified single solution method for the determination of phosphate in natural waters. Analytica Chimica Acta, 27, 31–36.
Nedelciu C.E., Ragnarsdottir K.V., Schlyter P., Stjernquist I. 2020. Global phosphorus supply chain dynamics: Assessing regional impact to 2050. Global Food Security, 26: 100426.
Neill M. 2005. A method to determine which nutrient is limiting for plant growth in estuarine waters—at any salinity. Marine Pollution Bulletin, 50, 945–955.
Neuwirth E. 2022. RColorBrewer: ColorBrewer palettes. R package version 1.1-3.
Otto S.B., Rall B.C., Brose U. 2007. Allometric degree distributions facilitate food-web stability. Nature, 450, 1226–1229.
Persson A., Hansson L., Brönmark C., Lundberg P., Pettersson L.B., Greenberg L., et al. 2001. Effects of enrichment on simple aquatic food webs. The American Naturalist, 157, 654–669.
R Core Team. 2023. R: a language and environment for statistical computing.
Rall B.C., Guill C., Brose U. 2008. Food-web connectance and predator interference dampen the paradox of enrichment. Oikos, 117, 202–213.
Redfield A. 1934. On the proportions of organic, derivatives in sea water and their relation to the composition of plankton, In: James Johnstone Memorial Volume. University Press of Liverpool, pp. 176–192.
Ringuet S., Sassano L., Johnson Z.I. 2011. A suite of microplate reader-based colorimetric methods to quantify ammonium, nitrate, orthophosphate and silicate concentrations for aquatic nutrient monitoring. Journal of Environmental Monitoring, 13, 370–376.
Ros M.B.H., Koopmans G.F., Van Groenigen K.J., Abalos D., Oenema O., Vos H.M.J., Van Groenigen J.W. 2020. Towards optimal use of phosphorus fertiliser. Scientific Reports, 10, 17804.
Rosenzweig M.L. 1971. Paradox of enrichment: destabilization of exploitation ecosystems in ecological time. Science, 171, 385–387.
Roy S., Chattopadhyay J. 2007. The stability of ecosystems: a brief overview of the paradox of enrichment. Journal of Biosciences, 32, 421–428.
Ryser R., Hirt M.R., Häussler J., Gravel D., Brose U. 2021. Landscape heterogeneity buffers biodiversity of simulated meta-food-webs under global change through rescue and drainage effects. Nature Communications, 12, 4716.
Shahmohamadloo R.S., Rudman S.M., Clare C.I., Westrick J.A., Wang X., Meester L.D., Fryxell J.M. 2022. Intraspecific genetic variation is critical to robust toxicological predictions of aquatic contaminants. Submitted to Nature Water (NATWATER-23-1190).
Shaw-McDonald S. 2019. Population responses to harvest selectivity are modified by temperature in an experimental population. University of Guelph.
Shaw N. 2020. Effects of glyphosate exposure, nutrient loading, and temperature on the population growth rates and cellular sizes of Chlorella vulgaris and Microcystis aeruginosa. University of Guelph.
Smithwick E.A.H. 2021. Nutrient flows in the landscape. In: The Routledge Handbook of Landscape Ecology. Routledge, London, pp. 140–158.
Tadiri C.P., Negrín Dastis J.O., Cristescu M.E., Gonzalez A., Fussmann G.F. 2024. Ecosystem connectivity and configuration can mediate instability at a distance in metaecosystems. Functional Ecology, 38, 153–164.
Takamura N., Otsuki A., Aizaki M., Nojiri Y. 1992. Phytoplankton species shift accompanied by transition from nitrogen dependence to phosphorus dependence of primary production in Lake Kasumigaura, Japan. Archiv für Hydrobiologie, 124, 129–148.
Tilman D. 1982. Resource competition and community structure: monographs in population biology. Princeton University Press, Princeton, N.J.
Tilman D. 1999. Global environmental impacts of agricultural expansion: the need for sustainable and efficient practices. Proceedings of the National Academy of Sciences, 96, 5995–6000.
Trainer V.L., Moore S.K., Hallegraeff G., Kudela R.M., Clement A., Mardones J.I., Cochlan W.P. 2020. Pelagic harmful algal blooms and climate change: lessons from nature's experiments with extremes. Harmful Algae, 91, 101591.
U.S. Environmental Protection Agency. 1993. Method 365.1: determination of phosphorus by semi-automated colorimetry (No. EPA/600/R-93/100). U.S. Environmental Protection Agency.
United Nations. 2022. World population prospects 2022: summary of results. United Nations, New York.
Veilleux B.G. 1979. An analysis of the predatory interaction between paramecium and didinium. The Journal of Animal Ecology, 48, 787.
Volterra V. 1926. Fluctuations in the abundance of a species considered Mathematically1. Nature, 118, 558–560.
Watkins J., Rudstam L., Holeck K. 2011. Length-weight regressions for zooplankton biomass calculations—A review and a suggestion for standard equations.
Wilke C.O., Wiernik B.M. 2022. ggtext: improved text rendering support for ’ggplot2. R package version 0.1.2.
Yadav M.R., Kumar R., Parihar C.M., Yadav R.K., Jat S.L., Ram H., et al. 2017. Strategies for improving nitrogen use efficiency: a review. Agricultural Reviews.
Yin X.W., Liu P.F., Zhu S.S., Chen X.X. 2010. Food selectivity of the herbivore daphnia magna (Cladocera) and its impact on competition outcome between two freshwater green algae. Hydrobiologia, 655, 15–23.
Zhang X., Zou T., Lassaletta L., Mueller N.D., Tubiello F.N., Lisk M.D., et al. 2021. Quantification of global and national nitrogen budgets for crop production. Nature Food, 2, 529–540.
Zou T., Zhang X., Davidson E.A. 2022. Global trends of cropland phosphorus use and sustainability challenges. Nature, 611,81–87.
Supplementary material
Supplementary Material 1 (PDF / 291 KB).
- Download
- 290.52 KB
Information & Authors
Information
Published In

FACETS
Volume 10 • 2025
Pages: 1 - 15
Editors: Josephine Gantois and Maud C.O. Ferrari
History
Received: 21 January 2024
Accepted: 27 November 2024
Version of record online: 13 March 2025
Notes
This paper is part of a collection entitled “Marginal land restoration, biodiversity conservation, and ecosystem services enhancement in agroecosystem landscapes”.
Copyright
© 2025 The Author(s). This work is licensed under a Creative Commons Attribution 4.0 International License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Data Availability Statement
The data and R scripts used for the analyses will be made publicly available on Github public repository upon publication (https://github.com/wangsh001/Nutrient_Transport).
Key Words
Sections
Subjects
Authors
Author Contributions
Conceptualization: XW, KSMC, RHH, JMF
Data curation: XW, MB, EMG
Formal analysis: XW, JMF
Funding acquisition: XW, KSMC, RHH, JMF
Investigation: XW, MB, EMG, RHH
Methodology: XW, MB, EMG, JMF
Project administration: XW
Resources: XW, KSMC, RHH, JMF
Software: XW, MB, EMG
Supervision: XW, KSMC, RHH, JMF
Validation: XW
Visualization: XW
Writing – original draft: XW
Writing – review & editing: XW, MB, EMG, KSMC, RHH, JMF
Competing Interests
The authors declare no competing interests.
Funding Information
University of Guelph Graduate Tuition Scholarship
Morwick Aquatic Biology Scholarship
This research was made possible through funding from the Food from Thought program through the Canada First Research Excellence Fund (CFREF), and NSERC Discovery Grant to JMF. XW received additional support through a Graduate Tuition Scholarship from the University of Guelph, Morwick Aquatic Biology scholarship, and an Ontario Graduate Scholarship from the Ontario government.
Metrics & Citations
Metrics
Other Metrics
Citations
Cite As
Xueqi Wang, Megan Braun, Ella McGuigan, Kevin S. McCann, Robert H. Hanner, and John M. Fryxell. 2025. The effect of nutrient transport downstream on food web stability in an experimental freshwater meta-ecosystem. FACETS.
10: 1-15.
https://doi.org/10.1139/facets-2024-0011
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
There are no citations for this item