Anthropogenic impacts on seagrass habitat connectivity: a model to explore potential links between human activity and marine invertebrate metapopulation persistence
Abstract
Coastal ecosystems face numerous stressors from anthropogenic activities. Furthermore, local stressors that directly impact ecological processes in one location can scale up to have indirect regional consequences. In the case of dispersing marine invertebrates, human activity may impact dispersal and survival rates, which can then alter connectivity patterns and metapopulation dynamics across the seascape. Here, we developed a framework to model metapopulation persistence of seagrass-associated invertebrates in the Salish Sea, focusing primarily on British Columbia, Canada. We combined a biophysical model of dispersal with metrics of habitat naturalness to model how local human activities that may impact dispersal can alter habitat connectivity patterns and potentially impact regional metapopulation persistence. We found that human activities that potentially impact dispersal can reduce population persistence across a region, although the effect varied based on location, population dynamics, and severity of the impacts modeled. The majority of populations, however, remained persistent, suggesting that there are robust and redundant pathways of dispersal that can maintain population connectivity in the face of local disturbances. This study highlights the importance of understanding human impacts and connectivity together in a regional context which could have implications for future marine spatial planning and the effective management of biodiversity.
1. Introduction
The connectivity of populations through dispersal is important for maintaining biodiversity in a region, especially following disturbances when ecological communities may be restructured and depleted (Thompson and Gonzalez 2017; Chase et al. 2020; Fontoura et al. 2022). Coastal marine habitats, in particular seagrass habitat and the species it supports, are under increasing threat from human disturbances such as coastal development, nutrient runoff, sedimentation, and the introduction of invasive species (Orth et al. 2017; Murphy et al. 2019; Turschwell et al. 2021). These stressors may impact a population's capacity to maintain local and regional patterns of diversity through increased mortality, reduced fitness, and altered dispersal ability (Hodgson et al. 2011; Jonsson et al. 2020). Despite the significance of these projected impacts, there is still a limited understanding of how human activities can alter connectivity patterns and influence biodiversity maintenance.
Population connectivity is the exchange of individuals between populations (Cowen and Sponaugle 2009), and in the case of a population being associated with a specific habitat type (e.g., seagrass), this connectivity can be made synonymous with habitat connectivity to emphasize the spatially discrete and structural component of the connectivity (Saura et al. 2014). Populations connected by dispersal in a region form metapopulations, thus linking regional processes to local species dynamics (Hanski 1998). Incorporating human impacts into the metapopulation concept can rescale our understanding of the spatial and temporal extent of impacts to seagrass (Carr et al. 2003; Gilarranz et al. 2017). An organism that disperses between two populations in seagrass habitat would experience stressors present in both locations. Therefore, a stressor that may only physically occur in one discrete seagrass habitat can then still have regional consequences by affecting the fitness of individuals immigrating and emigrating, thus altering the regional population dynamics of the linked metapopulation (Spromberg et al. 1998; Jonsson et al. 2020; Holstein et al. 2022). Human activities known to impact seagrass may also affect epifaunal invertebrate populations living within seagrass meadows given that measures of invertebrate abundance and diversity have been shown to scale with meadow fragmentation and shoot density (Lefcheck et al. 2016; Yeager et al. 2019). In addition, human activities are associated with changes in fish and invertebrate diversity (Iacarella et al. 2018). Knowing that there can be local impacts on both seagrass and animal diversity, and given the emerging recognition of the importance of connectivity to seagrass systems (Boström et al. 2010; Cristiani et al. 2021), it is possible that connectivity could spread impacts or confer population resilience in response to disturbances (Puritz and Toonen 2011; Jonsson et al. 2020).
Adequately conserving and managing biodiversity will require considering how connectivity and human impacts interact to influence population dynamics at multiple scales. This criterion is formalized in the UN Convention on Biological Diversity Aichi Target 11 qualitative elements for designing marine reserves, in which reserves should be “well-connected” and “integrated into the wider seascape”, among other criteria (UN CBD 2010). Integration implies that a reserve does not exist in isolation, and therefore it is important to consider potential cumulative impacts (CIs) existing in the region when designing a new marine reserve (Rees et al. 2018; Meehan et al. 2020). A CI framework quantifies the effect of multiple stressors on an ecosystem component, and it has been adapted to assess impacts at global (Halpern et al. 2008) and regional scales (Clarke Murray et al. 2015). At the local scale (e.g., one seagrass meadow), similar CI frameworks have been developed to identify potential impacts (Murphy et al. 2019; Nagel et al. 2020). In most CI frameworks, however, the possibility of remote stressors having indirect impacts in other locations through connectivity is not considered (Jonsson et al. 2020). This may be an important consideration for marine spatial planning, especially when areas of high naturalness are traditionally prioritized for protection (UN CBD 2008), but where it may be more important to protect an impacted area and limit stressors if it is also highly connected to other areas.
Here, we quantify the potential influence of human activity on the connectivity of seagrass-associated invertebrate populations in the Salish Sea, focusing primarily on British Columbia (BC), Canada with a minor contribution from Washington State, USA. Seagrass meadows form spatially discrete patches of habitat that can be used as a model system for studying connectivity across a seascape (Boström et al. 2006), and seagrass meadows in BC have been identified as ecologically and biologically significant areas (Rubidge et al. 2020). We assessed how the local stressors experienced by an animal population in a seagrass meadow may indirectly impact a population in another meadow that is connected by dispersal via deleterious effects on dispersing animals such that fewer individuals disperse or survive dispersal. To predict these regional impacts from local stressors, we combined connectivity and human activity data to model the persistence of regional metapopulations under varying levels of impacts. We hypothesized that metapopulations in regions with higher levels of human activity are more vulnerable because of the potential for impacts to dispersal to cascade among connected populations. This approach results in a framework that can project regional consequences of local stressors, and following validation, it can create new criteria for prioritizing important habitat with a combination of connectivity and naturalness characteristics.
2. Methods
To quantify the regional consequences of local stressors to seagrass habitat connectivity and metapopulation persistence, we first modeled the dispersal of invertebrates among seagrass habitat patches (Section 2.2) and quantified the multiple potential stressors affecting seagrass habitat (Section 2.3). With these measures, we then quantified the source and sink connectivity potential of each seagrass meadow in terms of the relative proportion of impacted invertebrates transferred to or received from other impacted meadows (i.e., regional impacts; Section 2.4). Lastly, we predicted the change to the persistence of regional metapopulations when human impacts alter connectivity (Section 2.5; Fig. 1).
Fig. 1.
2.1. Study area and system
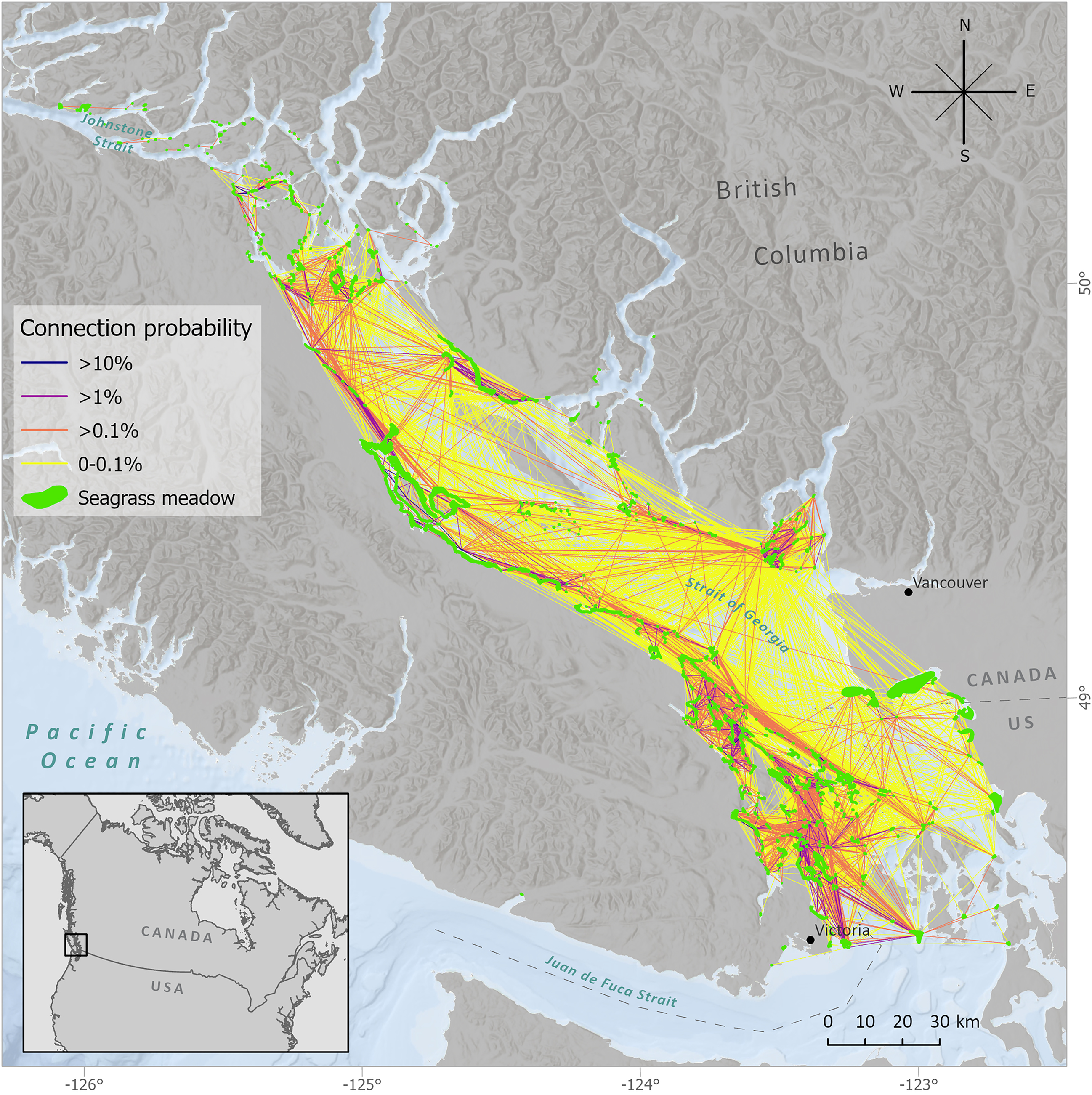
This study focused on seagrass meadows in the Salish Sea region of the BC and Washington coast in which eelgrass (Zostera marina) is the most abundant species but may co-occur with Zostera japonica. Eelgrass can occur to depths of ∼10 m (Christiaen et al. 2015), and meadows provide habitat for multiple trophic levels including a diverse epifaunal and infaunal invertebrate community (Heck et al. 2008; Huang et al. 2015; Knight et al. 2015; Whippo et al. 2018). We limited the study area to include only Canadian seagrass meadows and the few meadows in the United States that Canadian meadows are potentially connected to by invertebrate dispersal (selection criteria discussed in next section) covering an area of approximately 10 000 km2 (Fig. 2). The seagrass spatial dataset was compiled from multiple government and non-governmental sources (Cristiani et al. 2021). Most of the seagrass polygons were from the British Columbia Marine Conservation Analysis eelgrass dataset, which itself is a compilation of datasets from various sources using different survey methodologies (e.g., boat, ground, aerial) and collected over the past century (BCMCA 2010). Additional datasets were compiled from Washington State Department of Natural Resources (WSDNR 2018), Hakai Institute (Reshitnyk and Hakai Institute 2016), personal surveys, and restricted datasets from Fisheries and Oceans Canada and Islands Trust.
Fig. 2.
For measuring dispersal among meadows, we focused only on seagrass habitat specialists from the epifaunal invertebrate community (i.e., species that live exclusively in seagrass). As specialists, these species likely require direct dispersal connections between seagrass meadows for post-dispersal survival, however, many details of their life histories and dispersal strategies are unknown (Lefcheck et al. 2016). Specialist species considered include various gastropods (Lacuna variegata, Lottia alveus paralella, Phyllaplysia taylori) and the stalked jelly Haliclystus sanjuanensis (as identified in local field surveys from Stark et al. (2020) and Adamczyk (2022), and dispersal traits identified in Cristiani et al. (2021) and Forbes et al. (2025)). Dispersal for some of these species is possible through a planktonic larval dispersal stage, or in the case of direct developers, by rafting on buoyant detached seagrass or algal debris (Brooks and Bell 2001).
2.2. Modeling dispersal and connectivity
A key input for measuring metapopulation persistence is the probability of dispersal between meadows (i.e., spatially distinct patches) of seagrass habitat. We used dispersal probabilities obtained from an individual-based biophysical model in Cristiani et al. (2021). Briefly, this model used oceanographic currents from a hydrodynamic model (Soontiens et al. 2016; Olson et al. 2020) to simulate the passive dispersal of invertebrates among seagrass meadows in the Salish Sea. Individuals were tracked as they were advected and diffused across the seascape for up to 21 days. A daily mortality rate estimate of 15% day−1 was applied for all species. If an individual drifted over another seagrass meadow, it was considered settled and a successful connection was made between the origin and destination meadow. The probability of dispersal between two meadows was calculated as the number of individuals from the origin meadow settling on a destination meadow divided by the total number released from the origin meadow. These estimates represent the transport phase of connectivity (i.e., potential connectivity) and do not include modeling survival and reproduction in the destination seagrass meadow (i.e., realized connectivity). Simulations were ran across nine time periods to capture seasonal and yearly variation in dispersal, and connections were averaged across time to create one connectivity matrix c, containing the probability of connection among all combinations of meadows (see Cristiani et al. 2021 for additional model details).
We used a 21-day dispersal period to represent the planktonic drift duration trait of the various eelgrass specialists. For the species with planktonic larvae, their planktonic larval durations (PLDs) were equal to or greater than 21 days. Cristiani et al. (2021) found that there were minimal differences in dispersal patterns in the Salish Sea for PLDs greater than 21 days, and therefore the connectivity patterns generated by this dispersing period can likely be used to represent any species with dispersing durations greater than 21 days (see trait details in data repository for Cristiani et al. 2021). Also, in the case of species that may disperse by rafting (i.e., no pelagic larval phase), detached reproductive eelgrass shoots were found to be buoyant for up to 26 days (Harwell and Orth 2002; Källström et al. 2008).
We limited the study to Canadian meadows and the few US meadows connected to Canadian meadows due to limitations and inconsistencies of human activity data available for US meadows. The selection of US meadows we included was based on the topological characteristics of their connections to Canadian meadows (Fig. 2). We were only interested in meadows that may contribute to the persistence of a metapopulation of meadows (i.e., a source population), so we first retained any connections from a US meadow to a Canadian meadow. Of these US meadows, we then retained the connections between them (maintaining network “loops”) but did not include connections into these meadows from other US meadows that are not sources to Canadian meadows. These criteria resulted in retaining 32 US meadows, for a total of 685 meadows in the study.
2.3. Human activity and naturalness metrics
From spatial data, we quantified six anthropogenic activities/pressures that generate known stressors to seagrass, and by reducing habitat quality (e.g., reduced shoot growth and density, reduced water quality) these stressors may potentially impact the abundances and dispersal ability of invertebrate populations. For example, an activity (e.g., logging) may generate a stressor (e.g., increased sedimentation and reduced light), which may impact the system (e.g., reduced seagrass growth). Activities that potentially impact a meadow reduce the naturalness of the meadow—the relative level of human activity present in an area (UN CBD 2008; Rubidge et al. 2020). Our approach uses an existing coastal human impact model that was developed and applied in Canada (Murphy et al. 2019; Nagel et al. 2020). We adapted the model to include the marine and terrestrial activities for which data were available for the 685 meadows and their adjacent coastal watersheds across the region. A coastal watershed is the land area that channels water into a stream segment that drains into the ocean (termed “fundamental watershed” in the BC Freshwater Atlas; GeoBC 2021). The six direct and indirect pressures considered are: (1) human population in the adjacent watershed, (2) overwater structures within 1 km, (3) shoreline modification within 100 m of the adjacent shoreline, (4) agriculture in the adjacent watershed, (5) logging in the adjacent watershed, and (6) and presence of invasive European green crab (Carcinus maenas) within 1 km (Fig. 3).
Fig. 3.
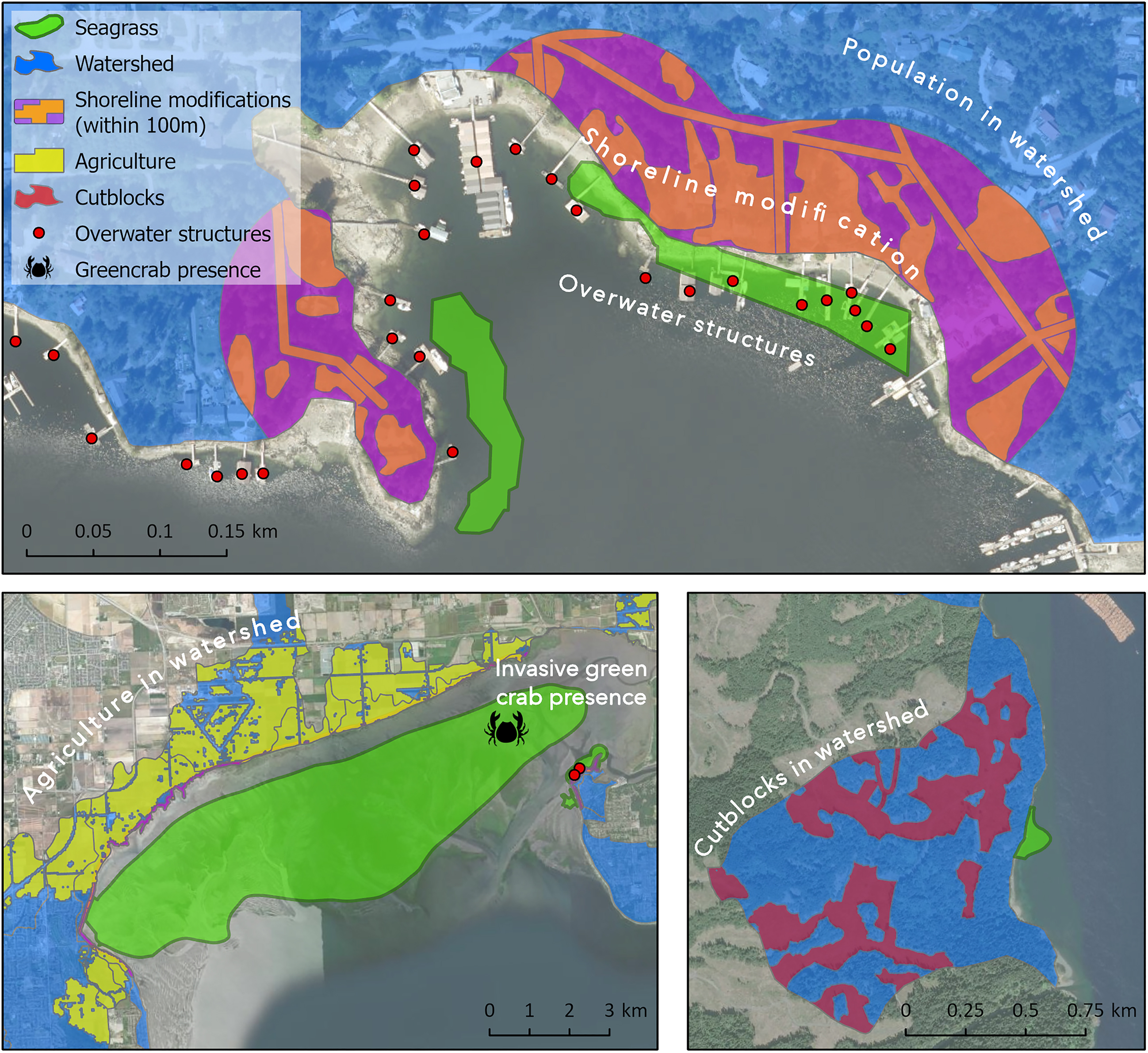
Human presence in the watershed may indicate increased levels of pollution and runoff as well as act as a proxy for additional activities not captured by the other metrics (e.g., recreation) (Shelton et al. 2016; Iacarella et al. 2018; Todd et al. 2019). Population counts (Center for International Earth Science Information Network 2017) were totaled for each watershed that is adjacent to a seagrass meadow. Only the human populations of watersheds bordering a seagrass meadow were associated to that meadow. Given that there are 685 meadows in our study area, many of which are in close proximity to each other, associating meadows only with their bordering watersheds allowed us to draw distinctions between meadows in terms of population counts. This is opposed to applying a large buffer around each meadow to capture any population stressors from distant watersheds that may decay with distance in the water.
Overwater structures (i.e., docks, float homes, marinas, aquaculture) can create shading over seagrass meadows and are indicative of increased boat traffic density with the potential for physical disturbance to seagrass and associated fauna (Burgin and Hardiman 2011; Skinner et al. 2013; Eriander et al. 2017; Iacarella et al. 2018). To quantify overwater structures, we calculated the area of overwater structures as a percentage of a 1 km buffer of water surface area around each seagrass meadow. We used an existing dataset (Iacarella et al. 2019) and added any additional overwater structures visible in recent Google Earth Imagery (∼2020) that were not present in the existing dataset.
Shoreline modification was quantified as the percentage of altered land within 100 m of the shoreline adjacent to the perimeter of each seagrass bed (Cristiani et al. 2023). Shoreline modifications (e.g., roads, buildings, ports, armoring, de-vegetated lawns, etc.) capture more direct localized pressures than watershed-level population measures. The 100 m distance was selected based on a suggested 30–100 m riparian buffer for sensitive marine habitat in Canada, such as eelgrass meadows (Lemieux et al. 2004). We used the most recent imagery available through Google Earth Pro to digitize modifications for all Canadian meadows (digitalization completed November 2020–February 2021). These modifications can increase runoff, pollution, and sedimentation, as well as alter wave dynamics and refraction patterns (Dethier et al. 2016; Quiros et al. 2017; Nahirnick et al. 2019; Todd et al. 2019).
Agricultural activities can increase sediment and nutrient loads entering the marine environment resulting in excessive algal growth and shading of the seagrass (Kemp et al. 2005; Uriarte et al. 2011). We calculated the percentage of agricultural land (Agriculture and Agri-Foods Canada 2021) occupying the watersheds adjacent to seagrass meadows. Coastal logging activities can also increase runoff and sediment load (Uriarte et al. 2011; Murphy et al. 2021). Using a dataset of cutblocks (i.e., harvested areas) in BC (BC Forest Analysis and Inventory Branch 2021), we calculated the percentage of cutblock areas active within the past 15 years occupying the watershed (the typical time for a forest stand to meet free-growing criteria; Forest Practices Board 2006). Lastly, we identified meadows within 1 km of any known occurrence of invasive European green crab (Howard and Therriault 2021). Green crabs are known to shred and consume eelgrass and significantly reduce meadow shoot density (Howard et al. 2019).
All metrics were normalized to 0–1 and compared using a principal components analysis. Metric vectors were mostly non-overlapping, confirming that this selection of metrics is adequate to use in our analysis for making distinctions between the large number of meadows (Fig. S1). For instance, logging activity is generally found in areas without significant human populations or other activities. Therefore, without this metric, meadows in the northern part of the Salish Sea would appear to have no activities present that may impact seagrass. Additionally, although human population and shoreline modification may be correlated, population is used to represent separate activities that may be occurring in the watershed compared to stressors associated with shoreline modification. Therefore, it is necessary to include both metrics in the overall impact score to emphasize the distinct impacts of heavily developed watersheds that would not be captured by the shoreline modification metric alone.
To achieve a relative comparison of impacts among meadows, we summed the normalized values of activities for each meadow. Data was compiled for only Canadian meadows (653 out of 685 meadows) because comparable human activity data was not available for the United States. The few directly connected meadows in the US were assigned the median value of impact as these were necessary to include to run the analysis. We used the median value of all meadows as opposed to the values of the closest Canadian meadows, as many connected US meadows were many kilometers away across Juan de Fuca Strait and we could not assume that they would have similar impact metrics (e.g., the less developed San Juan Islands, WA connected to meadows near the city of Victoria, BC). Lastly, we emphasize that our model of impacts is a simple additive model and is not considered a full CI assessment, which would require data on the vulnerability of eelgrass to each stressor (i.e., some activities may result in more impactful stressors than other activities). Therefore, we also refer to our additive metric of total impacts as a measurement of naturalness—the relative level of activity present in an area, with larger values being less natural (UN CBD 2008; Rubidge et al. 2020).
2.4. The regional distribution of impacts through dispersal
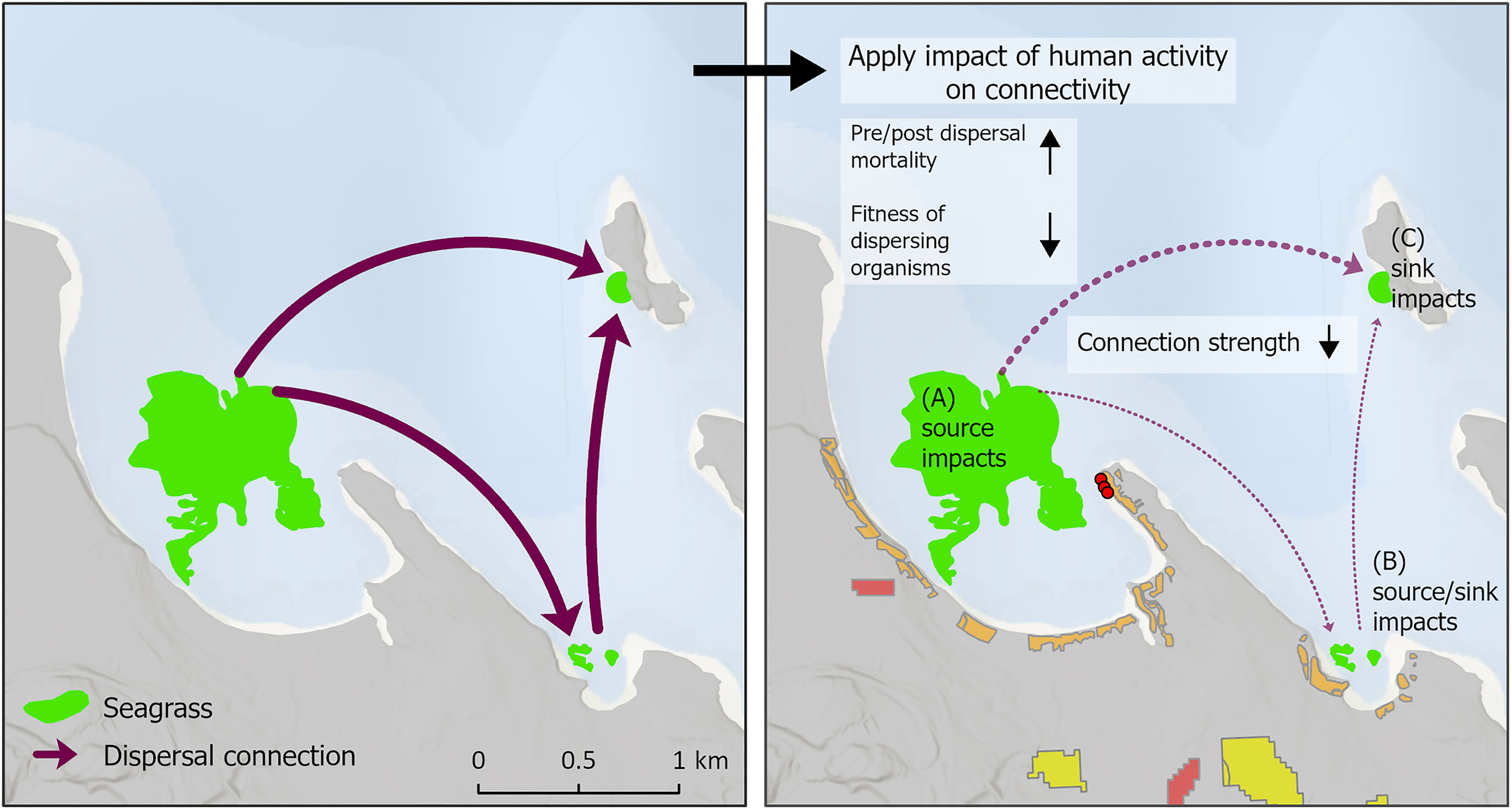
To assess how local impacts to habitat quality may indirectly impact invertebrate populations in other meadows via effects on animal movement, we summarized naturalness values based on the level of connectivity into and out of each meadow. Following an approach similar to Jonsson et al. (2020), we calculated a source- and sink-related impact metric for each seagrass meadow:where M is the total number of seagrass meadows, c is the connectivity matrix among all meadows, such that cij is the probability of dispersing between meadow i and meadow j (and i≠j), and S is the local naturalness of a meadow (i.e., the sum of the normalized activity values). In the case of the source impact, the dispersal connections leaving a meadow are multiplied by the local naturalness value (i), whereas with the sink impact the dispersal connections coming into a meadow are multiplied by the naturalness of each source meadow (j). This measure of source and sink impact captures the moderating effect human activities may have on dispersal success by increasing mortality and reducing fitness before and after dispersal such that fewer individuals disperse from the origin meadow, survive dispersal, or successfully reproduce in the destination meadow (Fig. 4). By assigning a value to each meadow, we can identify which meadows are both highly connected and impacted and therefore may cause the greatest disruption to the overall network of habitat connectivity.
(1)
(2)
Fig. 4.
2.5. Metapopulation persistence
To quantify the regional impact of human activities on habitat connectivity, we measured the change to metapopulation persistence when connectivity is altered by reduced naturalness. We adapted a density-dependent stochastic metapopulation model of persistence from Cristiani et al. (2024). Briefly, the model simulates population changes in individual meadows over multiple generations using the varying connectivity probabilities among meadows (Section 2.2). The population in a meadow at a given timestep (i.e., generation) is determined by the number of individuals immigrating/emigrating and the mortality rate of the adult population. In the case of marine invertebrates with planktonic larvae, all the individuals produced have a chance to disperse. Central to our model is the connectivity matrix (c) that determines the probability of dispersing between every pair of seagrass meadows. These probabilities are used to calculate the number of individuals successfully immigrating to another meadow. The model is initialized at the carrying capacity of the meadows, which we set to being proportional to the area of the meadow, such that the largest meadow started with 10 million individuals and the smallest meadow had 1000 individuals. These values were selected as illustrative examples intended to represent proportional differences in abundance among meadows but do not reflect actual abundances. We applied density dependence by reducing any population that was over carrying capacity at the end of each time step back to its carrying capacity. We used a 15% adult mortality rate and tested reproduction/emigration rates of 10%–60% of the population at each time step, such that all individuals produced will disperse but have a chance to recruit back to their home meadow. We ran our model for 1750 timesteps (i.e., generations), which was sufficient to reach a percentage of persistent populations that remained stable through time (see Cristiani et al. 2024 for model equations and additional details).
We then considered how the reduced naturalness of a source meadow may alter connectivity, in terms of the reduction in the transport of individuals, and potentially reduce persistence across the region. Building on the immigration equation in Cristiani et al. (2024) that calculates the probability of an individual arriving from another population:we modified how immigration is calculated to incorporate the potential reduction in the number of individuals emigrating from a source meadow. Immigration (Ii) is the total number of individuals coming into seagrass meadow i from all other meadows at time t:where M is the total number of seagrass meadows, c is the connectivity matrix among all meadows, such that cji is the probability of dispersing from meadow j to meadow i, Sjscaled is an adjusted local naturalness value of meadow j (described in the following paragraph), and Ej is the number of individuals dispersing from meadow j.
(3)
(4)
Since we do not know the relative vulnerability of seagrass to each activity or how stressors combine to potentially produce a cumulative effect, we compared five levels of naturalness to test different ranges of impact severity on metapopulation persistence. We scaled our total impact scores, Sj (which were normalized to a base level of 0–1), to the ranges of 1–2, 1–10, and 1–100 such that 1 is the most natural and the maximum value is the least natural. Scaled naturalness for meadow j is then calculated as:
(5)
Therefore, when dividing the dispersal probabilities in eq. 4 by these naturalness ranges, the probability of dispersal from the least natural meadow, for example, would be reduced by half (1–2 range), one order of magnitude (1–10 range), and two orders of magnitude (1–100 range), while the most natural meadows would be only minimally altered (i.e., divided by ∼1). We also ran a simulation without applying naturalness to use as a baseline comparison. To answer our question on the persistence of metapopulations influenced by human pressures, we calculated the percentage of populations that are persistent at the end of the simulations (i.e., 1750 generations), and we mapped the seagrass meadows that form persistent metapopulations for each naturalness range. For the map, we calculated the strongly connected components based on the connectivity among meadows. A component is a cluster of meadows that are connected to each other, and a strongly connected component is only the meadows of a component that can reach every other meadow in the component through directional connections. This excludes meadows that are only connected as sink populations that do not contribute back to the persistence of the metapopulation. This metric allows us to visualize the various roles that meadows play in the overall system (Artzy-Randrup and Stone 2010).
3. Results
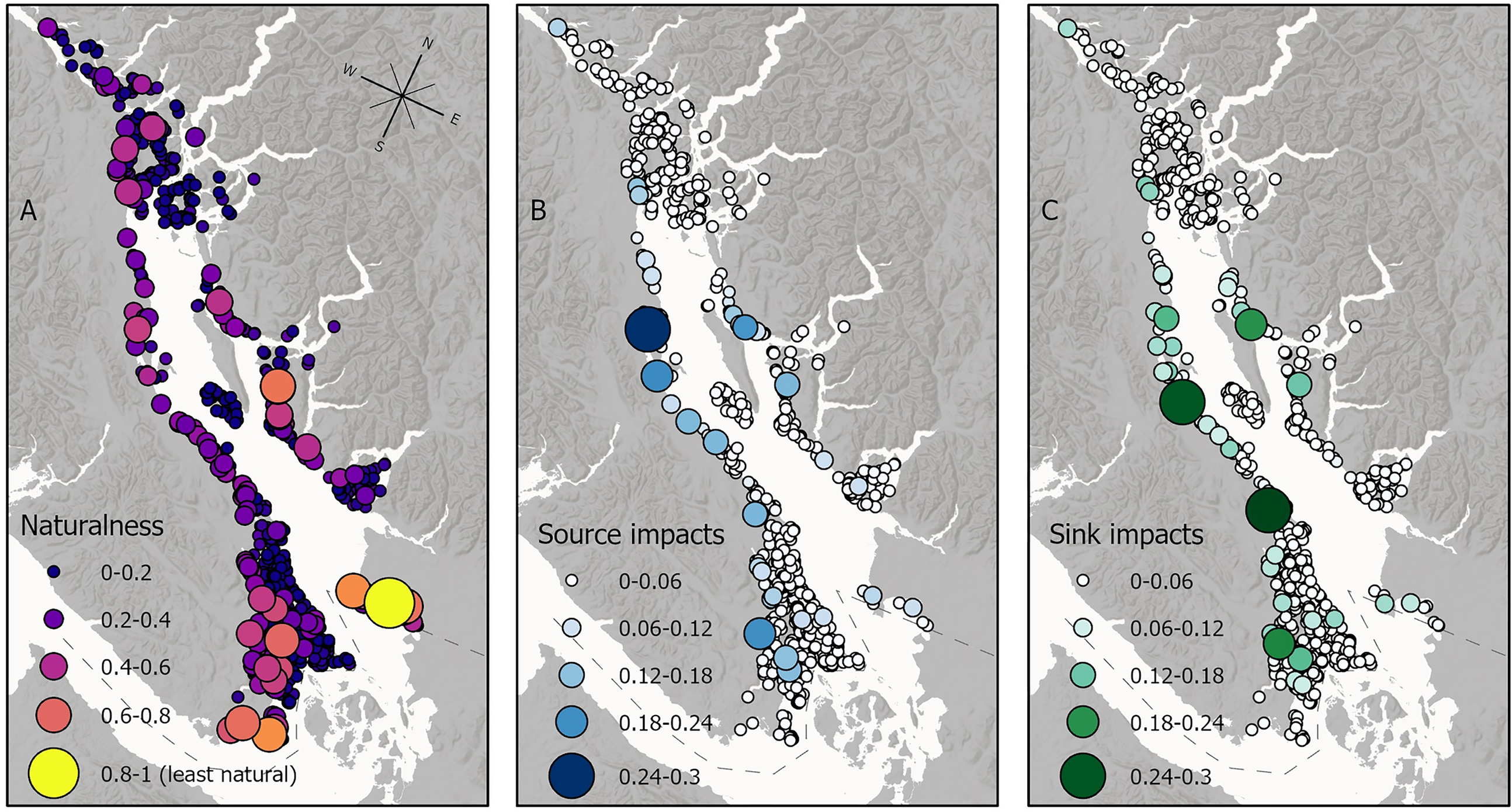
When assessing how the effects of local impacts on seagrasses and their associated faunal communities can transfer by altering animal dispersal, we found that areas of moderate to high naturalness can still experience or transfer the indirect effects of human activity due to patterns of connectivity across the seascape (Fig. 5). For example, the meadows that are the largest sources of impacts (Fig. 5B), are not necessarily the most locally impacted meadows (Fig. 5A)—these meadows are strongly connected to many other meadows and therefore have the potential to disrupt the overall network of habitat connectivity even if they only experience moderate local impacts. We also observed the opposite scenario, in which the least natural meadow (Boundary Bay, BC) is not a significant source or sink of impacts because it is not strongly connected to as many other meadows. Lastly, some meadows that have few local impacts, might still be receiving many impacted recruits because they are connected to highly impacted meadows as a sink population (Fig. 5C).
Fig. 5.
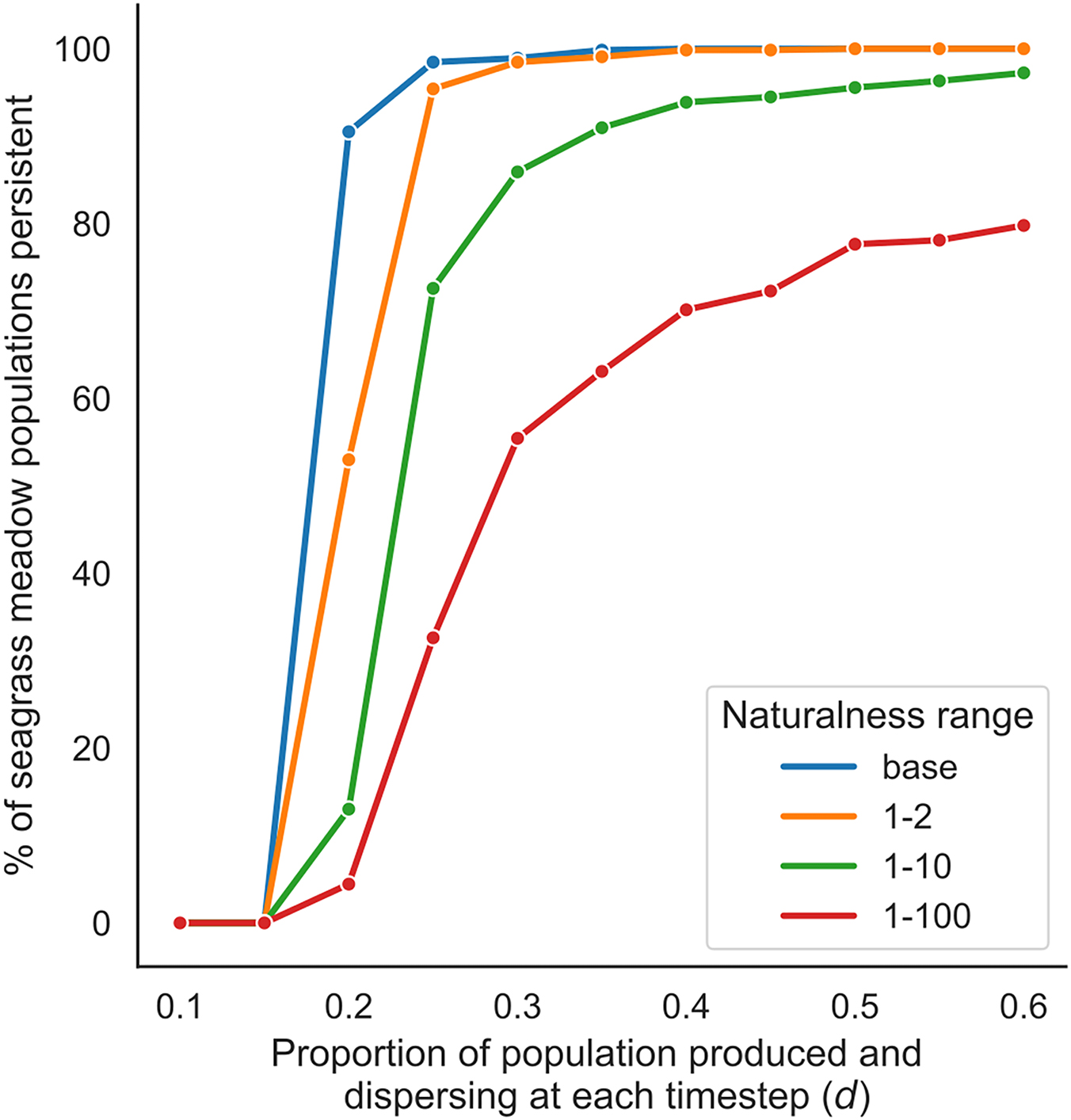
Building on the concept of sources and sinks of regionally transferred impacts, we then tested how changes to dispersal potential may impact metapopulation persistence. We found that human impacts to seagrass invertebrate populations can potentially reduce the number of persistent populations in a region due to reductions in the quantity and survival of dispersing individuals (Fig. 6). In the absence of regionally transferred impacts, invertebrate populations in nearly every meadow were persistent. We observed only minimal differences in persistence between the base level and 1–2 scaled range of naturalness. As human impacts intensified, the negative effects on regional population persistence also intensified (i.e., persistence was reduced by a larger amount at the 1–10 and 1–100 scaled ranges). The differences in persistence between ranges were greatest where the percentage of the population dispersing (d) was just above the mortality rate of the population (i.e., 15%). For example, at a dispersing rate of 20%, persistence ranged from 5% to 95% indicating that the level of impacts has the greatest effect at low levels of exchange.
Fig. 6.
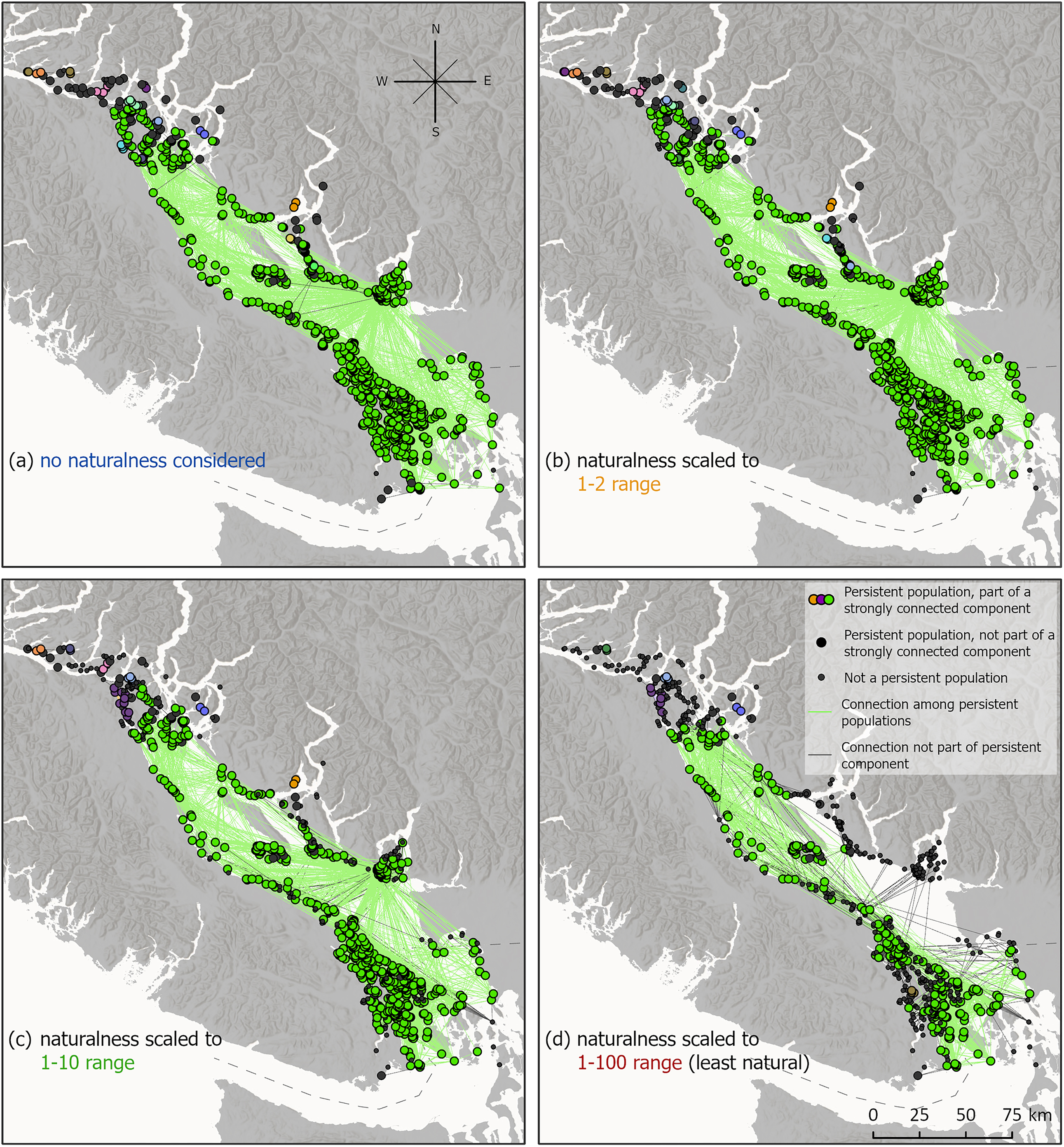
While we observed differences in persistence between ranges of naturalness, these differences were not consistent across the region. Due to numerous connection pathways, populations in many meadows were able to persist despite a reduction in recruits from some pathways. However, naturalness values were not evenly distributed in space—meadows in the southern part of the seascape were less natural—and therefore we observed more of a breakdown and splitting of the metapopulation network in these areas at the 1–10 and 1–100 naturalness ranges (Fig. 7).
Fig. 7.
4. Discussion
Using a metapopulation model that incorporates habitat connectivity and anthropogenic pressures, we found that local impacts may alter regional patterns of metapopulation persistence across the seascape for seagrass associated invertebrates. When we allowed impacts to influence connectivity, we saw a reduction in persistence; however, connectivity pathways and probabilities were redundant and robust enough to maintain persistence for most meadows except for a narrow range of invertebrate population values (i.e., when the percentage dispersing was only slightly greater than the population mortality rate). These patterns of persistence emphasize the importance of considering habitat as a network and the topological characteristics that emerge from the connectivity of the network.
A metapopulation model parameterized with dispersal connections that vary in magnitude and direction can uncover if there is redundancy of connectivity in the system that can overcome any reductions in dispersal from human activity (Minor and Urban 2007). This demographic “rescue effect” lowers the impact of local stressors and buffers regional biodiversity against local population size fluctuations (Thompson et al. 2016; Gilarranz et al. 2017; Harrison et al. 2020). By combining our metapopulation model with estimates of human impacts, we demonstrated a “safe operating space” of the network (Gonzalez et al. 2017), in which the persistence of the metapopulation was stable at low levels of impacts but was destabilized when the combination of impacts had a large cumulative effect. From these predictions, we can identify seagrass meadows that are both topologically important and highly impacted, where their loss would significantly reduce network connectivity and threaten overall metapopulation persistence. A key conservation challenge will then be to reconcile the priority to conserve the most natural meadows (UN CBD 2008; Rubidge et al. 2020) with the need to reduce impacts in and protect central but degraded meadows that potentially have greater regional influence.
While our model is largely conceptual, there is evidence of the effect of local and regional impacts on nearshore systems that support our findings. Cumulative effects from activities similar to those in this study, such as moorage and foreshore development, have been shown to reduce seagrass meadow area (Rees et al. 2023), CIs can influence invertebrate species assemblages among meadows (Adamczyk 2022), and nutrient inputs to seagrass meadows (similar to those from agriculture) can reduce invertebrate feeding rates (Tomas et al. 2015). Furthermore, Iacarella et al. (2018) found that the same suite of human pressures used in this study was correlated with a reduction in diversity of eelgrass associated fish communities in the Salish Sea. Assuming that these local pressures can impact other seagrass associated communities, we were then able to consider if these impacts are indirectly transferred and influence the dynamics of distant populations. For example, Jonsson et al. (2020) demonstrated that connectivity increased the CIs to blue mussel populations in the Baltic Sea by as much as 30% due to reduced larval production in source populations resulting in reduced recruitment opportunity in sink populations. Similar to our study, impacts from connectivity were not distributed equally across the seascape but were significant in specific areas. This pattern highlights the importance of considering connectivity for quantifying the full regional impact of human activities that otherwise would not be obvious if only considering impacts at their source (Jonsson et al. 2020). In another related study, point-source wastewater pollution in California was found to increase larval mortality and limit pelagic larval dispersal of sea stars. thus reducing connectivity and isolating populations of an otherwise high gene flow species (Puritz and Toonen 2011). Seagrass in the Salish Sea is predicted to be well-connected by invertebrate dispersal (Cristiani et al. 2021), however, as demonstrated in Puritz and Toonen (2011) and in our high CIs scenario, certain stressors can have significant impact on what is assumed to be a connected system.
Going forward, it will be important to validate and improve estimates for the three primary modeled components of the study: (1) connectivity, (2) metapopulation persistence, and (3) human impacts. Cristiani et al. (2021) discusses the technical limitations of the biophysical model, however, most fundamental is that our connectivity model only considers the reduction in potential connectivity, i.e., transport and settlement. The connectivity-impacts framework could be extended to understand the effects of human activity on realized connectivity, in which dispersing individuals successfully settle, survive, and reproduce in the sink meadow—creating a genetic connection (Cowen and Sponaugle 2009). Assessing this kind of connectivity would require measuring genetic structure in the Salish Sea, which could then be used to validate connectivity estimates from the biophysical model (Mertens et al. 2018; Wilcox et al. 2023).
To account for the uncertainty in the metapopulation model, we quantified persistence across a range of naturalness and population dynamic values. However, there are many areas of the modeling approach that can be refined to reduce uncertainty. Our results show that population persistence was not sensitive to moderate human activity if reproduction far exceeds mortality. When these rates were similar, however, persistence was much more sensitive to human impacts. This range of possible parameter values in the metapopulation model can be narrowed with species-specific empirical data, such as population mortality and reproduction rates obtained from experimental and observational studies. In addition, local impacts to the sink meadow could be incorporated into a more complex metapopulation model that applies meadow specific reproduction and mortality rates to further influence overall persistence through time. An approach that also incorporates the influence of species interactions and additional environmental conditions on fitness would then move us closer to a metacommunity model (Thompson et al. 2020).
A more accurate assessment of human impacts will require additional impact data, species-specific vulnerabilities to stressors, and knowledge of how stressors combine. Our model considers stressors from six activities, but additional activities at local, regional, and global scales are known to impact seagrass habitat (Agbayani et al. 2024; Murphy et al. 2024). For example, local activities like log storage can directly modify seagrass habitat, and regional development in the watershed, such as roads, can increase sedimentation (DFO 2023, 2024). Climate change indicators such as sea level rise, ocean temperature, acidification, and freshwater outflow temperature can impact seagrass (Weller et al. 2023; Murray et al. 2024) and temperature change can alter invertebrate dispersal (O'Connor et al. 2007; Gerber et al. 2014). Our impacts model could also be improved by incorporating a full CI assessment as opposed to a simple additive approach. This would first involve quantifying the vulnerability of seagrass and invertebrates to each stressor. For example, the impacts on seagrass from logging with proper precautions may be less than the impacts from agriculture in the watershed. This vulnerability of an ecosystem to stressors could be determined from qualitative expert-opinion scores (Teck et al. 2010) or from estimates of species’ responses to stressors (i.e., stressor-response curves), which provides an empirical relationship that can be validated with experimental and field observational studies (Rosenfeld et al. 2022; Jarvis et al. 2024). Lastly, we could improve our impacts model by understanding how stressors combine to have an effect greater than the sum of their parts (Halpern and Fujita 2013). For example, gradients of light and temperature have an interactive effect on eelgrass growth rate suggesting that multiple stressors can have nonlinear impacts (Dunic and Côté 2023).
We have demonstrated a framework that can be used to assess whether local human impacts influence the persistence of populations across a seascape by altering the quantity and survival of organisms dispersing among populations. This approach integrates two concepts which operate across scales: CIs and connectivity. Understanding these two concepts will be important for managing seagrass habitat and associated species in a landscape context, in which patterns of distribution, dispersal, and impacts will interact to influence regional management strategies (Murphy et al. 2021). Dispersal and the spread of impacts operate across borders, and it will be essential to coordinate research efforts to generate consistent datasets among Federal, Indigenous, Provincial and State governments. Although eelgrass is declining globally (Dunic et al. 2021), eelgrass in nearby Puget Sound, Washington is stable and resilient overall, despite a significant increase in local human and climatic stressors (Shelton et al. 2016). Assessing the relevance of managing for human impacts in the Salish Sea will therefore require a deeper understanding of seagrass and invertebrate responses to stressors and the mechanisms (e.g., dispersal) that allow for resilience to these stressors. Ultimately, refining and validating our models will increase their utility and promote their incorporation into broader marine spatial planning efforts.
References
Adamczyk E. 2022. Effects of human activities, environment, and time on eelgrass-associated epifaunal communities. University of British Columbia. Available from https://open.library.ubc.ca/collections/ubctheses/24/items/1.0417483.
Agbayani S., Nelson J.C., Schweitzer C., Vaidyanathan T., Murray C.C. 2024. Stability and reliability of regional cumulative impact mapping: a Canadian Pacific case study. Journal of Environmental Management, 371: 123105.
Agriculture and Agri-Foods Canada. 2021. Land Use 2010. Opend Data Canada. [accessed 1 November 2021].
Artzy-Randrup Y., Stone L. 2010. Connectivity, cycles, and persistence thresholds in metapopulation networks. PLoS Computational Biology, e1000876, 6(8).
BC Forest Analysis and Inventory Branch. 2021. Harvested areas of BC (Consolidated Cutblocks). BC Data Catalogue. Available from https://catalogue.data.gov.bc.ca/dataset/harvested-areas-of-bc-consolidated-cutblocks- [accessed 1 November 2021].
Boström C., Jackson E.L., Simenstad C. a. 2006. Seagrass landscapes and their effects on associated fauna: a review. Estuarine, Coastal and Shelf Science, 68(3–4): 383–403.
Boström C., Törnroos A., Bonsdorff E. 2010. Invertebrate dispersal and habitat heterogeneity: expression of biological traits in a seagrass landscape. Journal of Experimental Marine Biology and Ecology, 390(2): 106–117.
British Columbia Marine Conservation Analysis. 2010. Eelgrass Polygons. Available from https://bcmca.ca/data/eco_vascplants_eelgrass_polygons/ [accessed 12 November 2018].
Brooks R.A., Bell S.S. 2001. Mobile corridors in marine landscapes: enhancement of faunal exchange at seagrass/sand ecotones. Journal of Experimental Marine Biology and Ecology, 264(1): 67–84.
Burgin S., Hardiman N. 2011. The direct physical, chemical and biotic impacts on Australian coastal waters due to recreational boating. Biodiversity and Conservation, 20(4): 683–701.
Carr M.H., Neigel J.E., Estes J.A., Andelman S., Warner R.R., Largier J.L. 2003. Comparing marine and terrestrial ecosystems: implications for the design of coastal marine reserves. Ecological Applications, 13(1 SUPPL.): 90–107.
Center For International Earth Science Information Network-CIESIN-Columbia University. 2017. Gridded Population of the World, Version 4 (GPWv4): Population Density, Revision 11 (Version 4.11) [Data set]. Palisades, NY, Socioeconomic Data and Applications Center (SEDAC).
Chase J.M., Jeliazkov A., Ladouceur E., Viana D.S. 2020. Biodiversity conservation through the lens of metacommunity ecology. Annals of the New York Academy of Sciences, 1469(1): 86–104.
Christiaen B., Ferrier L., Dowty P., Gaeckle J., Berry H. 2015. Puget Sound Seagrass Monitoring Report. Washington Sate Department of Natural Resources. Available from https://sites.google.com/a/psemp.org/psemp/home.
Clarke Murray C., Agbayani S., Alidina H.M., Ban N.C. 2015. Advancing marine cumulative effects mapping: an update in Canada's Pacific waters. Marine Policy, 58: 71–77.
Cowen R.K., Sponaugle S. 2009. Larval dispersal and marine population connectivity. Annual Review of Marine Science, 1(1): 443–466.
Cristiani J., Rubidge E., Forbes C., Moore-Maley B., O'Connor M.I. 2021. A biophysical model and network analysis of invertebrate community dispersal reveals regional patterns of seagrass habitat connectivity. Frontiers in Marine Science, 8(August): 1–19.
Cristiani J., Rubidge E.M., Thompson P.L., Robb C.K., Hessing-Lewis M., O'Connor M.I. 2024. Quantifying marine larval dispersal to assess MPA network connectivity and inform future national and transboundary planning efforts. Canadian Journal of Fisheries and Aquatic Sciences, 81, 670–686.
Cristiani J.M., Bannar-Martin K.H., Rubidge E.M. 2023. Quantifying shoreline modifications adjacent to eelgrass meadows in the Strait of Georgia Bioregion. Canadian Technical Report of Fisheries and Aquatic Sciences, 3574: vi + 18.
Dethier M.N., Raymond W.W., McBride A.N., Toft J.D., Cordell J.R., Ogston A.S., et al. 2016. Multiscale impacts of armoring on Salish Sea shorelines: evidence for cumulative and threshold effects. Estuarine, Coastal and Shelf Science, 175: 106–117.
DFO. 2023. Coastwide evaluation and classification of Pacific Region estuaries based on anthropogenic activities and significant fish habitat. DFO Canadian Science Advisory Secretariat Science Advisory Report, 2023/039.
DFO. 2024. Geospatial indicators and metrics for threats to fish habitat in the Fraser River Basin with Thompson-Nicola as a case study. DFO Canadian Science Advisory Secretariat Science Advisory Report, 2024/041.
Dunic J.C., Côté I.M. 2023. Management thresholds shift under the influence of multiple stressors: eelgrass meadows as a case study. Conservation Letters, 16.
Dunic J.C., Brown C.J., Connolly R.M., Turschwell M.P., Côté I.M. 2021. Long-term declines and recovery of meadow area across the world's seagrass bioregions. Global Change Biology, 27, 4096–4109, Global Change Biology: gcb.15684.
Eriander L., Laas K., Bergström P., Gipperth L., Moksnes P.O. 2017. The effects of small-scale coastal development on the eelgrass (Zostera marina L.) distribution along the Swedish west coast—Ecological impact and legal challenges. Ocean and Coastal Management, 148: 182–194.
Esri. 2009. World Imagery. Available from https://www.arcgis.com/home/item.html?id=10df2279f9684e4a9f6a7f08febac2a9 [accessed 10 May 2021].
Esri. 2015. World Hillshade. Available from https://www.arcgis.com/home/item.html?id=1b243539f4514b6ba35e7d995890db1d [accessed 10 May 2021].
Esri. 2018. Ocean Basemap. Available from https://www.arcgis.com/home/item.html?id=67ab7f7c535c4687b6518e6d2343e8a2 [accessed 10 May 2021].
Fontoura L., D'Agata S., Gamoyo M., Barneche D.R., Luiz O.J., Madin E.M.P., et al. 2022. Protecting connectivity promotes successful biodiversity and fisheries conservation. Science, 375(6578): 336–340.
Forbes C., Hessing-Lewis M., O'Connor M.I. 2025. An experimental test of dispersal limitation of species diversity in eelgrass meadows. Estuaries and Coasts, 48.
Forest Practices Board. 2006. Achievement of free-growing forests–2004 provincial update. Forest Practices Board. Available from https://www.bcfpb.ca/wp-content/uploads/2016/04/SR25.pdf?itid=lk_inline_enhanced-template [accessed 1 November 2021].
GeoBC. 2021. Freshwater Atlas Watersheds. Available from https://catalogue.data.gov.bc.ca/dataset/freshwater-atlas-watersheds [accessed 1 November 2021].
Gerber L.R., Mancha-Cisneros M.D.M., O'Connor M.I., Selig E.R. 2014. Climate change impacts on connectivity in the ocean: implications for conservation. Ecosphere, 5(3): 1.
Gilarranz L.J., Rayfield B., Liñán-Cembrano G., Bascompte J., Gonzalez A. 2017. Effects of network modularity on the spread of perturbation impact in experimental metapopulations. Science, 357(6347): 199–201.
Gonzalez A., Thompson P., Loreau M. 2017. Spatial ecological networks: planning for sustainability in the long-term. Current Opinion in Environmental Sustainability, 29: 187–197.
Halpern B.S., Fujita R. 2013. Assumptions, challenges, and future directions in cumulative impact analysis. Ecosphere, 4(10): 1–11.
Halpern B.S., Walbridge S., Selkoe K.A., Kappel C. V., Micheli F., D'Agrosa C., et al. 2008. A global map of Human impact on marine ecosystems. Science, 319(February): 948–952.
Hanski I. 1998. Metapopulation dynamics. Trends in Ecology and Evolution, 4(6706): 113–114.
Harrison H.B., Bode M., Williamson D.H., Berumen M.L., Jones G.P. 2020. A connectivity portfolio effect stabilizes marine reserve performance. Proceedings of the National Academy of Sciences, 117(41): 25595–25600.
Harwell M.C., Orth R.J. 2002. Long-distance dispersal potential in a marine macrophyte. Ecology, 83(12): 3319–3330.
Heck K.L., Carruthers T.J.B., Duarte C.M., Randall Hughes A., Kendrick G., Orth R.J., Williams S.W. 2008. Trophic transfers from seagrass meadows subsidize diverse marine and terrestrial consumers. Ecosystems, 11(7): 1198–1210.
Hodgson J.A., Moilanen A., Wintle B.A., Thomas C.D. 2011. Habitat area, quality and connectivity: striking the balance for efficient conservation. Journal of Applied Ecology, 48(1): 148–152.
Holstein D.M., Smith T.B., van Hooidonk R., Paris C.B. 2022. Predicting coral metapopulation decline in a changing thermal environment. Coral Reefs, 41, 961–972.
Howard B.R., Therriault T.W. 2021. Ongoing range expansions of invasive marine invertebrates in the Pacific region. In State of the physical, biological and selected fishery resources of Pacific Canadian marine ecosystems in 2020. Canadian Technical Report of Fisheries and Aquatic Sciences, 3434. 203–207.
Howard B.R., Francis F.T., Côté I.M., Therriault T.W. 2019. Habitat alteration by invasive European green crab (Carcinus maenas) causes eelgrass loss in British Columbia, Canada. Biological Invasions, 21(12): 3607–3618.
Huang A.C., Essak M., O'Connor M.I. 2015. Top-down control by great blue herons Ardea herodias regulates seagrass-associated epifauna. Oikos, 124(11): 1492–1501.
Iacarella J.C., Adamczyk E., Bowen D., Chalifour L., Eger A., Heath W., et al. 2018. Anthropogenic disturbance homogenizes seagrass fish communities. Global Change Biology, 24(5): 1904–1918.
Iacarella J.C., Davidson I.C., Dunham A. 2019. Biotic exchange from movement of ‘static’ maritime structures. Biological Invasions, 21(4): 1131–1141.
Jarvis L., Rosenfeld J., Gonzalez-Espinosa P.C., Enders E.C. 2024. A process framework for integrating stressor-response functions into cumulative effects models. Science of the Total Environment, 906: 167456.
Jonsson P.R., Hammar L., Wåhlström I., Pålsson J., Hume D., Almroth-Rosell E., Mattsson M. 2020. Combining seascape connectivity with cumulative impact assessment in support of ecosystem-based marine spatial planning. Journal of Applied Ecology, 58(November): 576–586.
Källström B., Nyqvist A., Åberg P., Bodin M., André C. 2008. Seed rafting as a dispersal strategy for eelgrass (Zostera marina). Aquatic Botany, 88(2): 148–153.
Kemp W.M., Boynton W.R., Adolf J.E., Boesch D.F., Boicourt W.C., Brush G., et al. 2005. Eutrophication of Chesapeake Bay: historical trends and ecological interactions. Marine Ecology Progress Series, 303: 1–29.
Knight N.S., Prentice C., Tseng M., O'Connor M.I. 2015. A comparison of epifaunal invertebrate communities in native eelgrass Zostera marina and non-native zostera japonica at Tsawwassen, BC. Marine Biology Research, 11(6): 564–571.
Lefcheck J.S., Marion S.R., Orth R.J. 2016. Restored Eelgrass (Zostera marina L.) as a refuge for epifaunal biodiversity in Mid-Western Atlantic Coastal Bays. Estuaries and Coasts, 40, 200–212.
Lemieux J.P., Brennan J.S., Farrell M., Levings C.D., Myers D. 2004. Proceedings of the DFO/PSAT sponsored Marine Riparian Experts Workshop, Tsawwassen, BC, February 17-18, 2004. Can. Manuscr. Rep. Fish. Aquat. Sci. 2680: ix + 84p.
Meehan M.C., Ban N.C., Devillers R., Singh G.G., Claudet J. 2020. How far have we come? A review of MPA network performance indicators in reaching qualitative elements of Aichi Target 11. Conservation Letters, 13(August): 1–18.
Mertens L.E.A., Treml E.A., von der Heyden S. 2018. Genetic and biophysical models help define marine conservation focus areas. Frontiers in Marine Science, 5(August).
Minor E.S., Urban D.L. 2007. Graph theory as a proxy for spatially explicit populations models in conservation planning. Ecological Applications, 17(6): 1771–1782.
Murphy G.E.P., Dunic J.C., Adamczyk E.M., Bittick S.J., Côté I.M., Cristiani J., et al. 2021. From coast to coast to coast : ecology and management of seagrass ecosystems across Canada. Facets, 6: 139–179.
Murphy G.E.P., Stock A., Kelly N.E. 2024. From land to deep sea: A continuum of cumulative human impacts on marine habitats in Atlantic Canada. Ecosphere, 15(9): e4964.
Murphy G.E.P., Wong M.C., Lotze H.K. 2019. A human impact metric for coastal ecosystems with application to seagrass beds in Atlantic Canada. Facets, 4(1): 210–237.
Murray C.C., Kelly N.E., Nelson J.C., Murphy G.E.P., Agbayani S. 2024. Cumulative impact mapping and vulnerability of Canadian Marine ecosystems to anthropogenic activities and stressors. DFO Can. Sci. Advis. Sec. Res. Doc. 2024/024: vi + 46.
Nagel E.J., Murphy G.E.P., Fast J., Bittick S.J., Adamczyk M., Connor M.I.O., et al. 2020. Application of a coastal human impact metric and nitrogen loading model to 10 eelgrass (Zostera marina) meadows in British Columbia. Canadian Technical Report of Fisheries and Aquatic Sciences, 3429: vii + 45p.
Nahirnick N.K., Costa M., Schroeder S., Sharma T. 2019. Long-term eelgrass habitat change and associated Human impacts on the west coast of Canada. Journal of Coastal Research, 36(1): 30.
O'Connor M.I., Bruno J.F., Gaines S.D., Halpern B.S., Lester S.E., Kinlan B.P., Weiss J.M. 2007. Temperature control of larval dispersal and the implications for marine ecology, evolution, and conservation. Proceedings of the National Academy of Sciences, 104(4): 1266–1271.
Olson E.M., Allen S.E., Do V., Dunphy M., Ianson D. 2020. Assessment of nutrient supply by a tidal jet in the Northern Strait of Georgia based on a biogeochemical model. Journal of Geophysical Research: Oceans, 125(8): 1–25.
Orth R.J., Carruthers T.I.M.J.B., Dennison W.C., Duarte C.M., James W., Jr K.L.H., et al. 2017. A global crisis for seagrass ecosystems. Bioscience, 56(12): 987–996.
Puritz J.B., Toonen R.J. 2011. Coastal pollution limits pelagic larval dispersal. Nature Communications, 2(1).
Quiros T.E.A.L., Croll D., Tershy B., Fortes M.D., Raimondi P. 2017. Land use is a better predictor of tropical seagrass condition than marine protection. Biological Conservation, 209: 454–463.
Rees M.J., Knott N.A., Astles K.L., Swadling D.S., West G.J., Ferguson A.M., et al. 2023. Cumulative effects of multiple stressors impact an endangered seagrass population and fish communities. Science of the Total Environment, 904: 166706.
Rees S.E., Foster N.L., Langmead O., Pittman S., Johnson D.E. 2018. Defining the qualitative elements of Aichi Biodiversity Target 11 with regard to the marine and coastal environment in order to strengthen global efforts for marine biodiversity conservation outlined in the United Nations Sustainable Development Goal 14. Marine Policy, 93(October 2016): 241–250.
Reshitnyk L., Hakai Institute. 2016. Eelgrass extent—coastal British Columbia. Available from https://catalogue.hakai.org/dataset/ca-cioos_0219e1d6-8dfc-4718-b89b-ea3dff06a70d [accessed 12 November 2018].
Rosenfeld J., Gonzalez-Espinosa P., Jarvis L., Enders E., Bayly M., Paul A., et al. 2022. Stressor-response functions as a generalizable model for context dependence. Trends in Ecology and Evolution, 37(12): 1032–1035.
Rubidge E., Jeffery S., Gregr E.J., Gale K.S.P., Frid A. 2020. Assessment of nearshore features in the Northern Shelf Bioregion against criteria for determining ecologically and biologically significant areas (EBSAs). DFO Can. Sci. Advis. Sec. Res. Doc. 2020/023: vii + 63p.
Saura S., Bodin Ö., Fortin M.J. 2014. Editor's choice: stepping stones are crucial for species’ long-distance dispersal and range expansion through habitat networks. Journal of Applied Ecology, 51(1): 171–182.
Shelton A.O., Francis T.B., Feist B.E., Williams G.D., Lindquist A., Levin P. 2016. Forty years of seagrass population stability and resilience in an urbanizing estuary. Journal of Ecology, 105, 458–470.
Skinner M.A., Courtenay S.C., McKindsey C.W. 2013. Reductions in distribution, photosynthesis, and productivity of eelgrass Zostera marina associated with oyster Crassostrea virginica aquaculture. Marine Ecology Progress Series, 486: 105–119.
Soontiens N., Allen S.E., Latornell D., Le Souëf K., Machuca I., Paquin J.-P., et al. 2016. Storm surges in the Strait of Georgia simulated with a regional model. Atmosphere-Ocean, 54(1): 1–21.
Spromberg J.A., John B.M., Landis W.G. 1998. Metapopulation dynamics: indirect effects and multiple distinct outcomes in ecological risk assessment. Environmental Toxicology and Chemistry, 17(8): 1640–1649.
Stark K., Thompson P., Yakimishyn J., Lee L., Adamczyk E., Hessing-Lewis M., O'Connor M. 2020. Beyond a single patch: local and regional processes explain diversity patterns in a seagrass epifaunal metacommunity. Marine Ecology Progress Series, 655: 91–106.
Teck S., Halpern B., Kappel C. 2010. Using expert judgement to estimate marine ecosystem vulnerability in the California Current. Ecological Applications, 20(5): 1402–1416.
Thompson P.L., Gonzalez A. 2017. Dispersal governs the reorganization of ecological networks under environmental change. Nature Ecology and Evolution, 1(6).
Thompson P.L., Guzman L.M., De Meester L., Horváth Z., Ptacnik R., Vanschoenwinkel B., et al. 2020. A process-based metacommunity framework linking local and regional scale community ecology. Ecology Letters, 23(9): 1314–1329.
Thompson P.L., Rayfield B., Gonzalez A. 2016. Loss of habitat and connectivity erodes species diversity, ecosystem functioning, and stability in metacommunity networks. Ecography, 40(October 2016): 98–108.
Todd P.A., Heery E.C., Loke L.H.L., Thurstan R.H., Kotze D.J., Swan C. 2019. Towards an urban marine ecology: characterizing the drivers, patterns and processes of marine ecosystems in coastal cities. Oikos, 128, 1215–1242.
Tomas F., Martínez-Crego B., Hernán G., Santos R. 2015. Responses of seagrass to anthropogenic and natural disturbances do not equally translate to its consumers. Global Change Biology, 21(11): 4021–4030.
Turschwell M.P., Connolly R.M., Dunic J.C., Sievers M., Buelow C.A., Pearson R.M., et al. 2021. Anthropogenic pressures and life history predict trajectories of seagrass meadow extent at a global scale. Proceedings of the National Academy of Sciences, 118(45): e2110802118.
UN CBD. 2008. Decision adopted by the conference of the parties to the convention on biological diversity at its ninth meeting. UNEP/CBD/COP/DEC/IX/20. [accessed 6 December 2021].
UN CBD. 2010. Strategic plan for biodiversity 2011–2020: provisional technical rationale, possible indicators and suggested milestones for the Aichi biodiversity targets. UNEP/CBD/COP/10/27/Add.1. [accessed 6 December 2021].
Uriarte M., Yackulic C.B., Lim Y., Arce-Nazario J.A. 2011. Influence of land use on water quality in a tropical landscape: a multi-scale analysis. Landscape Ecology, 26(8): 1151–1164.
Washington State Department of Natural Resources. 2018. Nearshore habitat eelgrass monitoring. Available from https://www.dnr.wa.gov/programs-and-services/aquatics/aquatic-science/nearshore-habitat-eelgrass-monitoring [accessed 12 November 2018].
Weller J.D., Moore R.D. (Dan), Iacarella J.C. 2023. Stream thermalscape scenarios for British Columbia, Canada. Canadian Water Resources Journal / Revue canadienne des ressources hydriques, 49, 233–252.
Whippo R., Knight N.S., Prentice C., Cristiani J., Siegle M.R., O'Connor M.I. 2018. Epifaunal diversity patterns within and among seagrass meadows suggest landscape-scale biodiversity processes. Ecosphere, 9(11): e02490.
Wilcox M.A., Jeffery N.W., DiBacco C., Bradbury I.R., Lowen B., Wang Z., et al. 2023. Integrating seascape resistances and gene flow to produce area-based metrics of functional connectivity for marine conservation planning. Landscape Ecology, 38, 2189–2205.
Yeager L.A., Geyer J.K., Fodrie F.J. 2019. Trait sensitivities to seagrass fragmentation across spatial scales shape benthic community structure. Journal of Animal Ecology, 88(11): 1743–1754.
Supplementary material
Supplementary Material 1 (PDF / 116 KB).
- Download
- 116.98 KB
Information & Authors
Information
Published In

FACETS
Volume 10 • 2025
Pages: 1 - 15
Editors: David Lesbarrères and Yangfan Zhang
History
Received: 24 July 2024
Accepted: 2 February 2025
Version of record online: 20 March 2025
Copyright
© 2025 Authors Cristiani, O'Connor, and The Crown. This work is licensed under a Creative Commons Attribution 4.0 International License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Data Availability Statement
The code and result datasets generated for this study are available at https://doi.org/10.5281/zenodo.14829603.
Key Words
Sections
Subjects
Authors
Author Contributions
Conceptualization: JC, EMR, MIO
Data curation: JC
Formal analysis: JC
Funding acquisition: EMR, MIO
Methodology: JC
Supervision: EMR, MIO
Visualization: JC
Writing – original draft: JC, EMR, MIO
Writing – review & editing: JC, EMR, MIO
Competing Interests
The authors declare there are no competing interests.
Funding Information
This research was sponsored by the NSERC Canadian Healthy Oceans Network and its Partners: Department of Fisheries and Oceans Canada and INREST (representing the Port of Sept-Îles and City of Sept-Îles). It was also funded through the Fisheries and Oceans Canada Strategic Program for Ecosystem-based Research and Assessment (SPERA).
Metrics & Citations
Metrics
Other Metrics
Citations
Cite As
John Cristiani, Emily M. Rubidge, and Mary I. O'Connor. 2025. Anthropogenic impacts on seagrass habitat connectivity: a model to explore potential links between human activity and marine invertebrate metapopulation persistence. FACETS.
10: 1-15.
https://doi.org/10.1139/facets-2024-0152
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
There are no citations for this item