Winter diet of five sympatric ungulates in west-central Alberta, Canada—inference from DNA metabarcoding of fecal pellets
Abstract
Diet is one of the determinants of ungulate ecology. However, there have been few studies of the diet of sympatric ungulates in western Canada. We used DNA metabarcoding of fecal samples to evaluate the winter diet of white-tailed deer (Odocoileus virginianus), mule deer (O. hemionus), moose (Alces americanus), elk (Cervus canadensis), and caribou (Rangifer tarandus caribou) in west-central Alberta. We found that forbs comprised 53%–82% of the reads from plant DNA, while mosses, sedges, grasses, and shrubs made up relatively little of each ungulate’s diet. There was considerable overlap in the winter diet of the five ungulates (Horn-Morisita index = 42%–64% overlap; Bray-Curtis index = 20%–45% overlap). Moose had the most distinct diet, while elk and caribou had the least distinct. However, our analysis was restricted to a coarse taxonomic resolution (family). Lichen species were not identifiable, beyond their fungal component and a number of taxonomic assignments were difficult to rationalise given our knowledge of the diet of the five ungulate species and the ecology of west-central Alberta, Canada. DNA metabarcoding of fecal samples is efficient, but apparent limitations with the technique restrict the description and quantification of diet of the five species of ungulate that we studied.
Introduction
Climate and landscape change are altering the boreal forest at unprecedented rates (Pimm et al. 1993; Sala et al. 2000). Climate plays a significant role in determining vegetation growth rate (Brecka et al. 2018), plant distribution (Soja et al. 2007), and phenological timelines (Post et al. 2018; Park et al. 2020). Concurrently, the boreal forest is experiencing rapid rates of deforestation, primarily due to fire and forestry, but also fossil fuel extraction, surface mining, and agriculture (Hansen et al. 2013; Pickell et al. 2015). These disturbances convert mature forest into early-seral stands, altering the composition and productivity of the plant community (Haeussler et al. 2002; Sulla-Menashe et al. 2018). The effects of climate and landscape change have cascading impacts, including on herbivores who rely on these changing plant communities for nutrition (Bradshaw et al. 2003; Weisberg and Bugmann 2003). For example, the transition of mature forest to early-seral stands typically increases forage availability for ungulates during the summer (Strong and Gates 2006; Edenius et al. 2015; Finnegan et al. 2019), although the effect varies according to silviculture treatments such as planting (Boan et al. 2011), herbicide application (Strong and Gates 2006; Koetke et al. 2023), and removal or burning of woody material that remains after harvest (Edenius et al. 2014).
Nutritional ecology examines how an organism’s forage requirements and acquisition are influenced by ecological factors such as food quantity and quality, community interactions, and environmental conditions (Raubenheimer et al. 2009). Nutrition is dictated by forage quality and availability, and affects individual behavior, including movement and distribution (Hebblewhite et al. 2008; Bischof et al. 2012; Tattersall et al. 2023), as well as recruitment and survival (Monteith et al. 2013; Fisher et al. 2020; Jackson et al. 2021). For example, Proffitt et al. (2016) found that elk (Cervus canadensis) with access to lower quality nutrition during spring had poorer body condition during the following winter, resulting in reduced pregnancy rates.
Diet, or the ingestion of nutrients, is inherently important to understanding an animal’s nutritional ecology, including outcomes for reproduction and survival. However, it is difficult to quantify diet for many free-ranging species. Typical methods are expensive, invasive, or time-intensive (e.g., Hodder et al. 2013; Thompson et al. 2015; Denryter et al. 2017). Recent advancements in DNA metabarcoding have improved our ability to quantify diet items in pellet samples (Pompanon et al. 2012; Ando et al. 2020). DNA metabarcoding uses PCR amplification and high-throughput sequencing to identify the genetic signatures of plants and fungi (de Sousa et al. 2019). In comparison to microhistology, metabarcoding can be more cost effective (Nichols et al. 2015), less labour intensive (King and Schoenecker 2019; Littleford-Colquhoun et al. 2022), and can reveal greater taxonomic precision (Port et al. 2016). However, there is still some uncertainty in establishing the relationship between quantifiable DNA fragments (i.e., read counts) and the biomass of the diet item that was consumed. There also are questions about sample age and viability and potential challenges associated with the resolution of the reference library when differentiating genetically similar diet items (Bonin et al. 2020).
Woodland caribou (Rangifer tarandus caribou), an ungulate found across Canada, are forage specialists during winter that feed primarily on terrestrial and arboreal lichens (Thomas et al. 1996). In comparison, moose (Alces americanus), deer (Odocoileus spp.), and elk, sympatric ungulates found across much of the range of caribou, are more generalist foragers (Franzmann and Schwartz 1997). Moose consume over 200 species, although willow (Salix spp.) and birch (Betula spp.) are common forage items across much of the North American range of the species (Franzmann and Schwartz 1997). In Alberta, the winter diet of moose is dominated by trees and shrubs (Nowlin 1978; Renecker and Hudson 1986, 1988). To the best of our knowledge, the diet of white-tailed deer (Odocoileus virginianus) has not yet been evaluated in western North America. In other regions, white-tailed deer are known to have a varied diet including deciduous trees and shrubs, coniferous trees, forbs, grasses, and fruits (Hewitt 2011). During winter, mule deer consume true firs (Abies spp.) and Douglas fir (Pseudotsuga menziesii), deciduous shrubs, and various forbs (Williams et al. 1980; Hodder et al. 2013; Rea et al. 2017). In general, elk consume grasses (Holsworth 1960; Woods 1972; Salter and Hudson 1980; Churchill 1982; Kohl et al. 2012), however, forbs, shrubs, sedges, and ground litter also are common diet items (Holsworth 1960; Woods 1972; Salter and Hudson 1980; Churchill 1982). A few studies found trace amounts of conifers and mosses in the winter diet of elk (Woods 1972; Churchill 1982; Kohl et al. 2012).
Despite the importance of forage for an animal’s ecology, there are few recent studies of the diet of sympatric ungulates in western Canada. We used metabarcoding of pellet samples to identify the winter diets of deer, moose, elk, and caribou found across west-central Alberta. Similarities and differences among diets indicate potential niche overlap that may lead to competitive exclusion. This is particularly significant for understanding the predator-prey dynamics of woodland caribou, an Endangered species in much of western Canada, and for understanding apparent competition (DeCesare et al. 2010), where forage subsidies are hypothesized to be attracting other prey species into caribou ranges (Fisher and Burton 2021; Fuller et al. 2023).
We used Proportion of Occurrence (POO; Deagle et al. 2019) and Relative Read Abundance (RRA; Deagle et al. 2019) to assess differences in the composition of winter diet among the five species of ungulate. We expected the diet of caribou to consist mostly of lichen (Thomas et al. 1996). Moose and elk typically forage on willow and birch, and grass, respectively (Holsworth 1960; Woods 1972; Nowlin 1978; Salter and Hudson 1980; Churchill 1982; Renecker and Hudson 1986, 1988; Kohl et al. 2012). The diet of mule and white-tailed deer has not been evaluated in Alberta, but based on research from other regions we expected deer to present as true generalists with diets consisting of a variety of plants and fungi in equal proportions (Franzmann and Schwartz 1997; Hewitt 2011).
Methods
Study area
The study area covers approximately 21 000 km2 across west-central Alberta, in Treaty 8 territory that includes the traditional lands of the Aseniwuche Winewak, Dane-zaa, Métis, Nehiyawak, Simpcw, Stoney and Tsuut’ina peoples (Fig. 1). Highway 16 serves as the approximate southern boundary, with the study area extending north towards Grande Prairie, Alberta. The southwest portion of the study area is primarily mountainous, with minimal anthropogenic disturbance. The northeast portion of the area extends into the boreal forest and is licensed for forestry and oil and gas development. Anthropogenic disturbances, including cutblocks, roads, pipelines, seismic lines, and well sites, are prevalent. The primary natural disturbance in this region is wildfire.
Fig. 1.
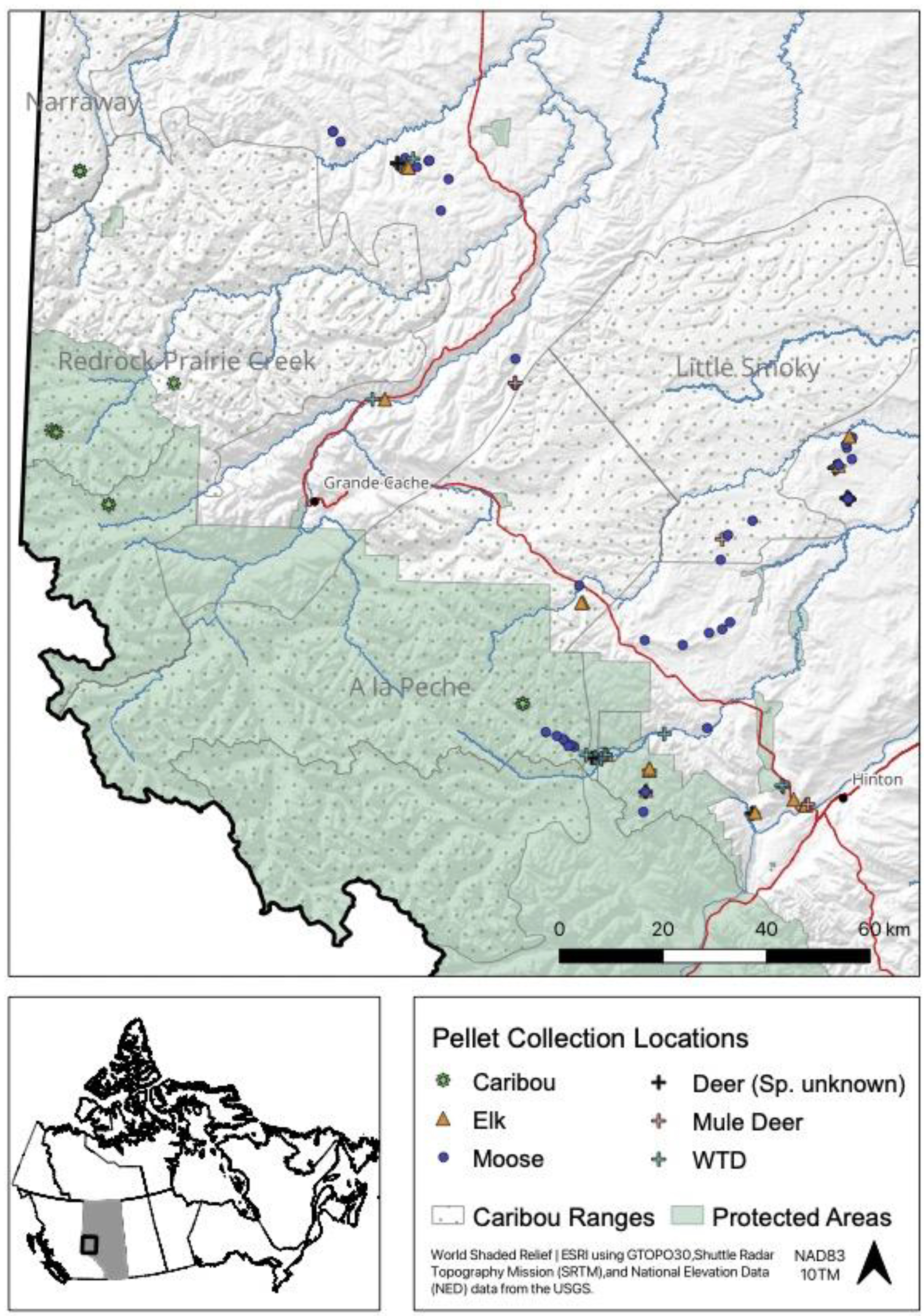
The mountainous portions of the study area include alpine, subalpine, and montane natural subregions (Natural Regions Committee 2006). The alpine subregion is largely non-vegetated with herbaceous meadows and shrub cover. The subalpine and montane subregions are primarily mixed forests, with lodgepole pine (Pinus contorta) and Engelmann spruce (Picea engelmannii) dominant in the subalpine, and lodgepole pine, Douglas fir (Pseudotsuga menziesii), white spruce (Picea glauca), and aspen (Populus tremuloides) dominant in the montane subregion. The boreal forest includes the Upper and Lower Foothills natural subregions (Natural Regions Committee 2006). Both subregions are heavily forested, with the Lower Foothills including aspen, lodgepole pine, and white spruce, and the Upper Foothills lodgepole pine, black spruce, and white spruce. The study area has a diverse mammalian community. Ungulate species include moose, white-tailed deer, mule deer, elk, mountain goats (Oreamnos americanus), bighorn sheep (Ovis canadensis), and caribou. Populations of woodland caribou (Narraway, Redrock-Prairie Creek, A la Peche, and Little Smoky) were assessed as Threatened or Endangered by the Committee on the Status of Endangered Wildlife in Canada (COSEWIC 2014).
Pellet sampling
We collected deer, moose, and elk pellets in west-central Alberta from January to April of 2022. The study area was stratified by forest cover type according to the Alberta Satellite Land Cover dataset (Alberta Agriculture and Forestry 2017). We randomly selected ten locations using QGIS Random Selection tool from each forest cover type in each region and used those locations as potential sampling sites. We also collected samples opportunistically by following fresh ungulate tracks when encountered in the field. Typically, we collected samples by searching a designated area 72–96 h after snowfall. Some samples were collected from fresh roadkill carcasses. All spatial analyses were completed using QGIS tools and software (QGIS Development Team 2021).
Samples, consisting of 10–25 pellets, were collected from one pellet group, and only one sample was collected for each set of tracks. We froze samples immediately after collection and they were kept frozen at −20 °C until processing. We used a number of indicators or direct measures to identify the ungulate that deposited the pellets. This included an inspection of the pellet, according to size and shape; identification of tracks in the snow, that presumably matched the individual species that deposited the pellets; and assessment of habitat relative to the ecology of the focal ungulate species. We carried out pellet collection in Willmore Wilderness Park, Rock Lake Provincial Park, and Rock Lake–Solomon Creek Wildland in conjunction with existing Government of Alberta activities and in accordance with Research and Collection Permit No: 22-039. Caribou pellets were collected in the winter of 2016 by visiting areas used by GPS-collared caribou (Permit No: 16-008). Although there was a 6-year gap between sampling for caribou and the pellets of deer, elk, and moose, the diet of low-elevation mountain caribou was thought to be relatively simple during the winter (Thomas et al. 1996; Johnson et al. 2001; Webber et al. 2022). Furthermore, most caribou samples were taken from protected areas with considerably less industrial activity that may have influenced the composition of plant and lichen communities as well as the distribution and behaviour of caribou (e.g., Dyer et al. 2001; Johnson et al. 2015). Pellet morphology is similar for white-tailed deer and mule deer, thus, we used genetic analysis to identify each deer sample to species (Bach et al. 2022).
DNA extraction and amplification
We prepared pellet samples for barcoding analysis by homogenizing approximately 45 mLs of pellets with 3–5 mLs of 95% ethanol. All samples were sent to the Canadian Center for DNA Barcoding at the University of Guelph for analysis of plant and fungal DNA. In the lab, DNA was further homogenized and extracted from the supplied samples using established methods (Ivanova et al. 2006). DNA sequences were amplified using PCR with the following primer cocktails: ITS-S2F_t1/ITS3_t1/ITS4_t1 and ITS-Bry3d_t1/ITS-S2F-Bry_t1/ITSBry4d_t1/ITS4d_t1. Sequence reads were compared to the Barcode of Life Data System (BOLD reference library; www.boldsystems.org) and assigned a taxonomic identifier (see Moran et al. 2019 for detailed explanation of method). Separately, unidentified pellets from white-tailed or mule deer were analyzed for nuclear microsatellite DNA at the INRA11 loci and mitochondrial 16S rRNA gene by Wildlife Genetics International. We prepared samples for genetic testing by swabbing the extremity of partially thawed samples with porous toothpicks. At the lab, DNA was purified using QIAGEN DNeasy Blood and Tissue kits (Qiagen, Hilden, Germany).
Data analysis
We considered taxa that returned >100 reads per sample as valid (Paula et al. 2022). Most pellet samples resulted in plant or fungal identification at a relatively coarse taxonomic resolution (e.g., genus or family), thus, we summarized results to taxonomic family. We identified probable lichenicolous fungi using www.lichenicolous.net (Lawrey and Diederich 2018). We used the Alberta Conservation Management Information System (ACMIS) databases for lichen and fungi (Government of Alberta 2022) and Lichens of North America (Brodo et al. 2001) to cross-reference lichen that were found in our study area.
Statistical analysis
We conducted two analyses of the identified plant and fungal material for each pellet sample by ungulate species. First, we used Proportion of Occurrence (POO; Deagle et al. 2019) to evaluate the presence of plant and fungal family in all samples for each ungulate. We calculated POO as the proportion of fecal samples that contained (i.e., presence) a particular diet item. The Frequency of Occurrence (FOO) was normalized so that the proportion occurrence of the full set of diet items summed to 100%. We used the Horn-Morisita similarity index (Horn 1966) to compare the POO of each sample and compared samples among ungulates, with 1.0 representing complete overlap between diets.
Second, we used the Relative Read Abundance (RRA; Deagle et al. 2019) to infer the relative abundance of plant and fungal families within each sample. We calculated RRA as the number of reads for each taxon divided by the total number of reads for that sample. Those data were then summarised for all samples for each ungulate species. We used the Bray–Curtis similarity index (Bray and Curtis 1957) to compare the RRA between samples, with 1.0 representing complete overlap. In theory, both POO and RRA should produce similar trends in diet among the five ungulate species that we sampled, but the two measures differ in how the underlying data are represented. Occurrence metrics (e.g., POO) represent presence/absence of taxa in a sample according to a threshold in the number of sequences (i.e., conversion of sequence counts to a single occurrence of a diet item) and RRA represents the proportional summaries of sequence counts of each diet item. POO and RRA can have unique biases and are both common in the literature, thus, our choice to summarise the sequence data using both metrics (Deagle et al. 2019).
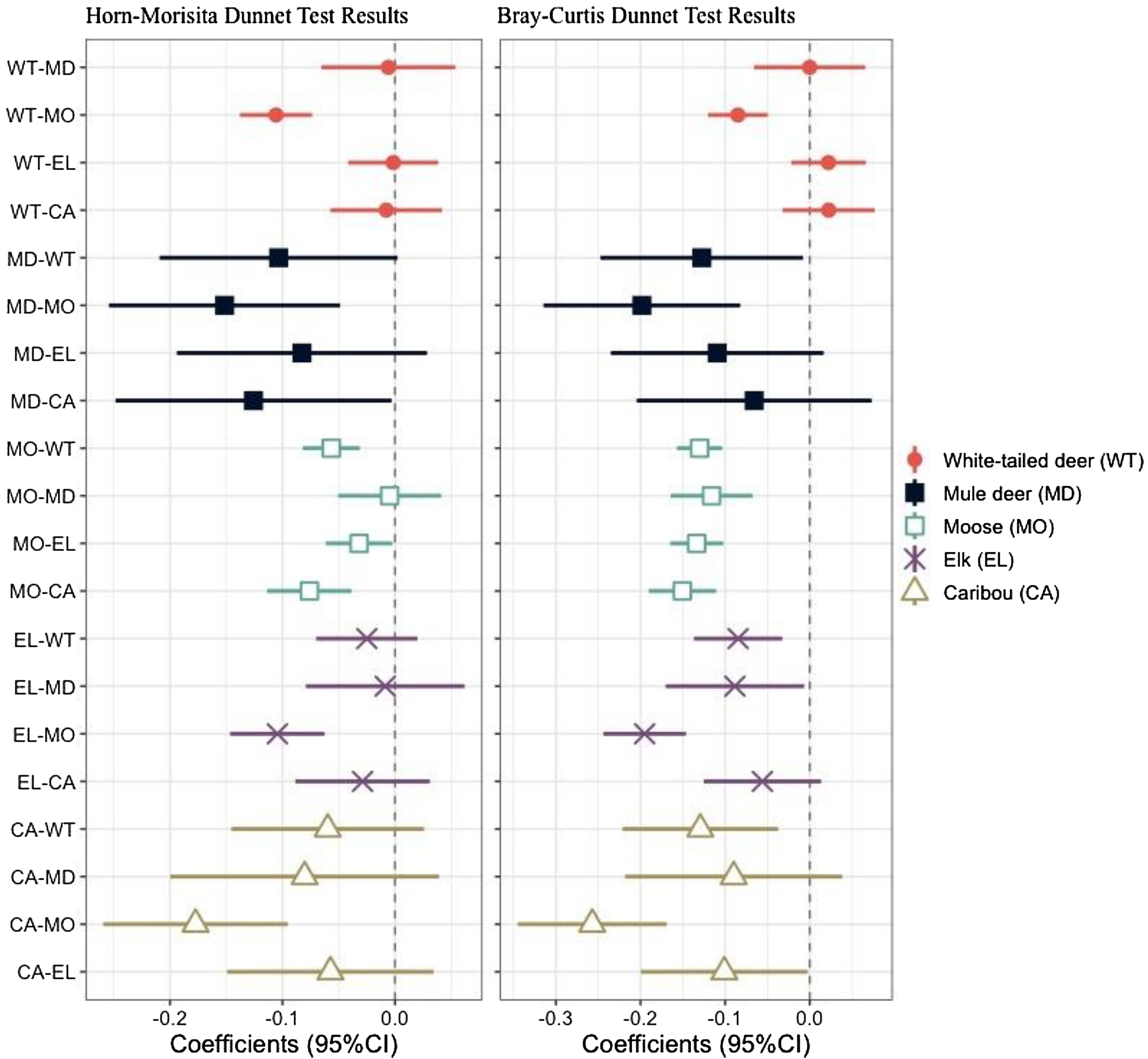
We conducted a pairwise comparison of the Horn-Morisita and Bray-Curtis indices for each sample and calculated the mean similarity among samples. We evaluated the data for normality by plotting residuals and by visually inspecting Q–Q plots. We then used an Analysis of variance (ANOVA) (ANOVA) followed by a Dunnett’s test to evaluate significant differences in the mean similarity indices for each ungulate. We used R (v4.1.3; R Core Team 2022) and the Vegan package (Oksanen et al. 2022) to conduct the statistical analyses.
Results
We submitted 88 samples for metabarcoding analysis that were collected from 88 discrete sites (Supplemental Table 1). In most cases, sampled pellet groups were >500 m apart (N = 46), however, some groups were within 100 m (N = 14) of another pellet group, but from a discrete or unique trail in the snow. We used genetic analyses to identify the deer species of 28 of 29 pellet samples. The source of one sample from a white-tailed deer was visually identified. The final dataset included 24 white-tailed deer (systematic sample = 4, opportunistic sample = 20), 5 mule deer (systematic = 3, opportunistic = 2), 36 moose (systematic = 10, opportunistic = 26), 15 elk (systematic = 2, opportunistic = 13), and 8 caribou samples (systematic = 0, opportunistic = 8). There were approximately 6.9 million reads from the 88 samples. From the metabarcoding analysis, there were 3.7 million reads for plant DNA and 3.1 million reads for fungal DNA. In total, 921 123 reads were unidentified or unknown at the family level. On average, each submitted sample returned 43 587 (range: 204–195 469) reads of plant DNA and 35 737 (range: 0–195 206) reads of fungal DNA. Samples from white-tailed deer comprised 33% of the total reads, while 8% were from mule deer, 38% from moose, 14% from elk, and 6% from caribou. These values were roughly in-line with the proportion of samples submitted for each ungulate.
Diet composition
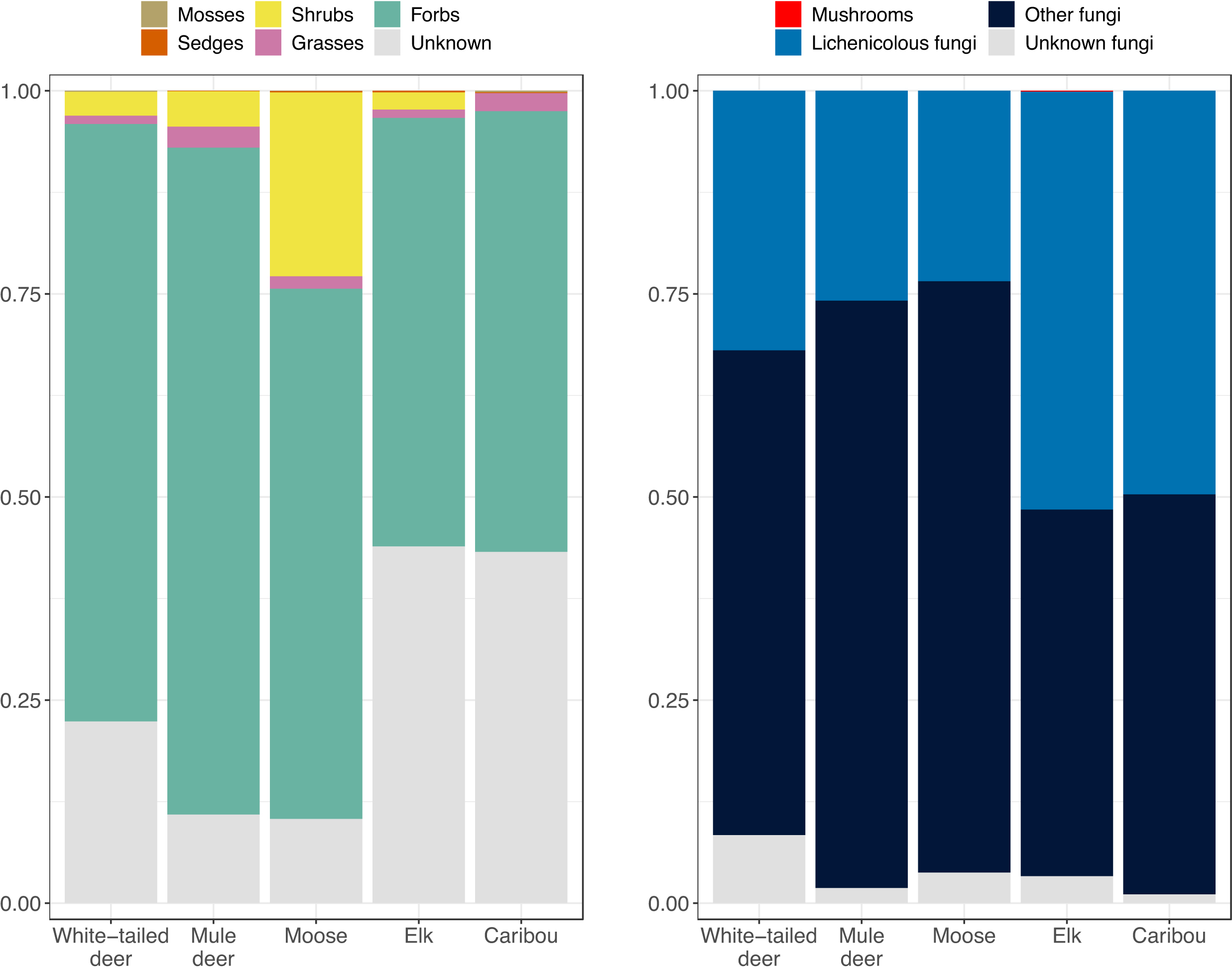
Thirty-seven distinct taxonomic families were identified by the metabarcoding process (Fig. 2; Supplemental Table 2). Bionectriaceae, Herpotrichiellaceae, Nectriaceae, Pleosporaceae, Pleosporaceae incertae sedis, and Tremellaceae were families of fungi classified as “probably lichenicolous” and associated with a lichen species found in our study area (Brodo et al. 2001; Lawrey and Diederich 2018; Government of Alberta 2022). For all five ungulate species, Lentibulariaceae (carnivorous plants, common butterworts) was one of the most frequent plant families identified within the winter pellet samples (>75% of all samples). Poaceae (grass) was identified in >70% of samples for white-tailed deer, mule deer, and caribou. Grimmiaceae and Santalaceae (mosses and sandlewoods) were only found in samples identified as white-tailed deer, Andreaeaceae and Melanthiaceae (mosses and bunchflower) were found only in caribou samples, and Salicaceae (willow) was found only in moose samples.
Fig. 2.
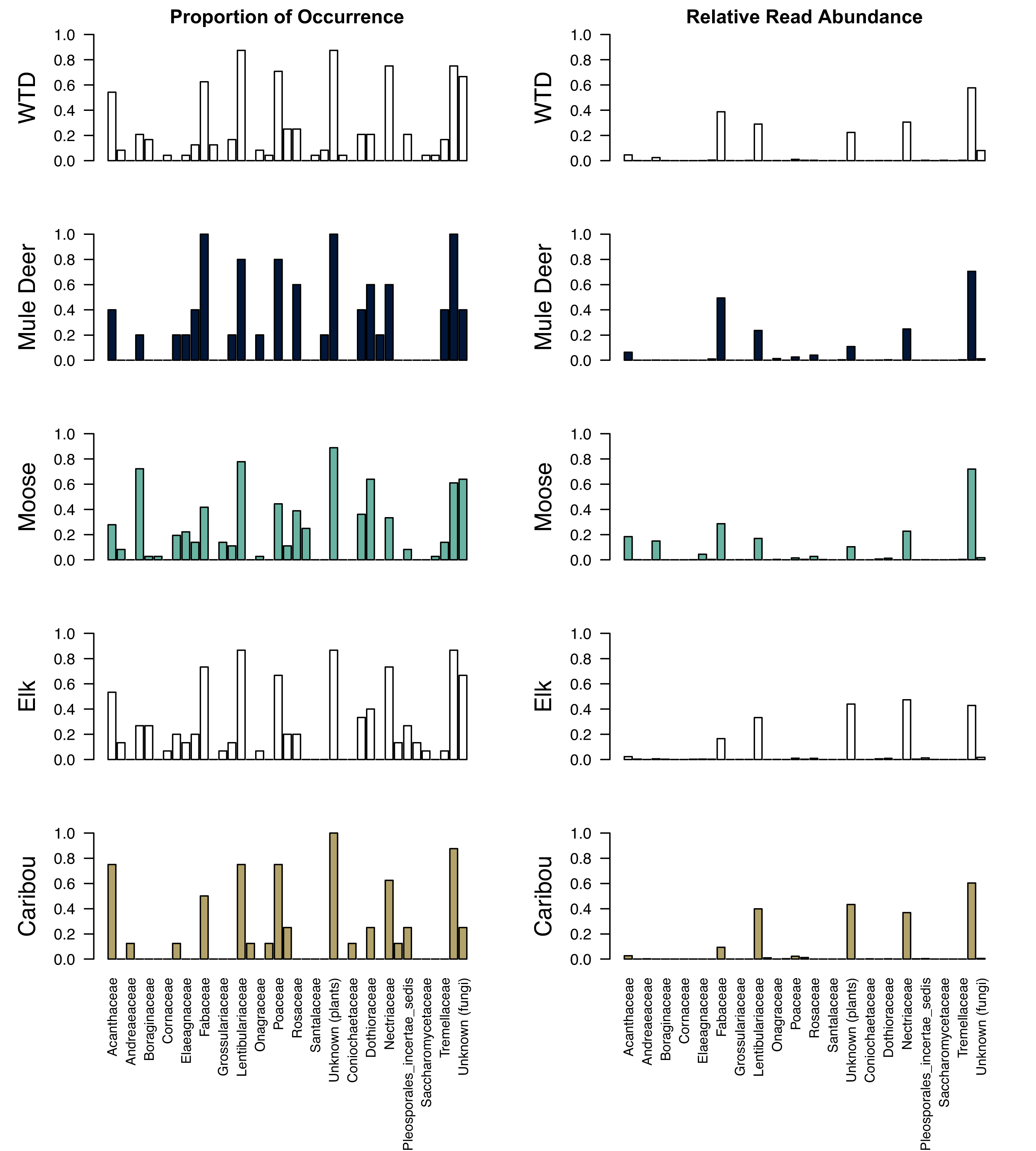
The most frequently identified fungal family was associated with decaying organic matter (Trichocomaceae). Nectriaceae, a lichen-associated fungi, was identified in >70% of samples for white-tailed deer and elk. Bionectriaceae, another lichen associated fungi, was only found in white-tailed deer samples, and Herpotrichiellaceae, also lichen associated fungi, was only found in mule deer samples. Psathyrellaceae, a common mushroom fungus in our study area, was only found in elk pellet samples.
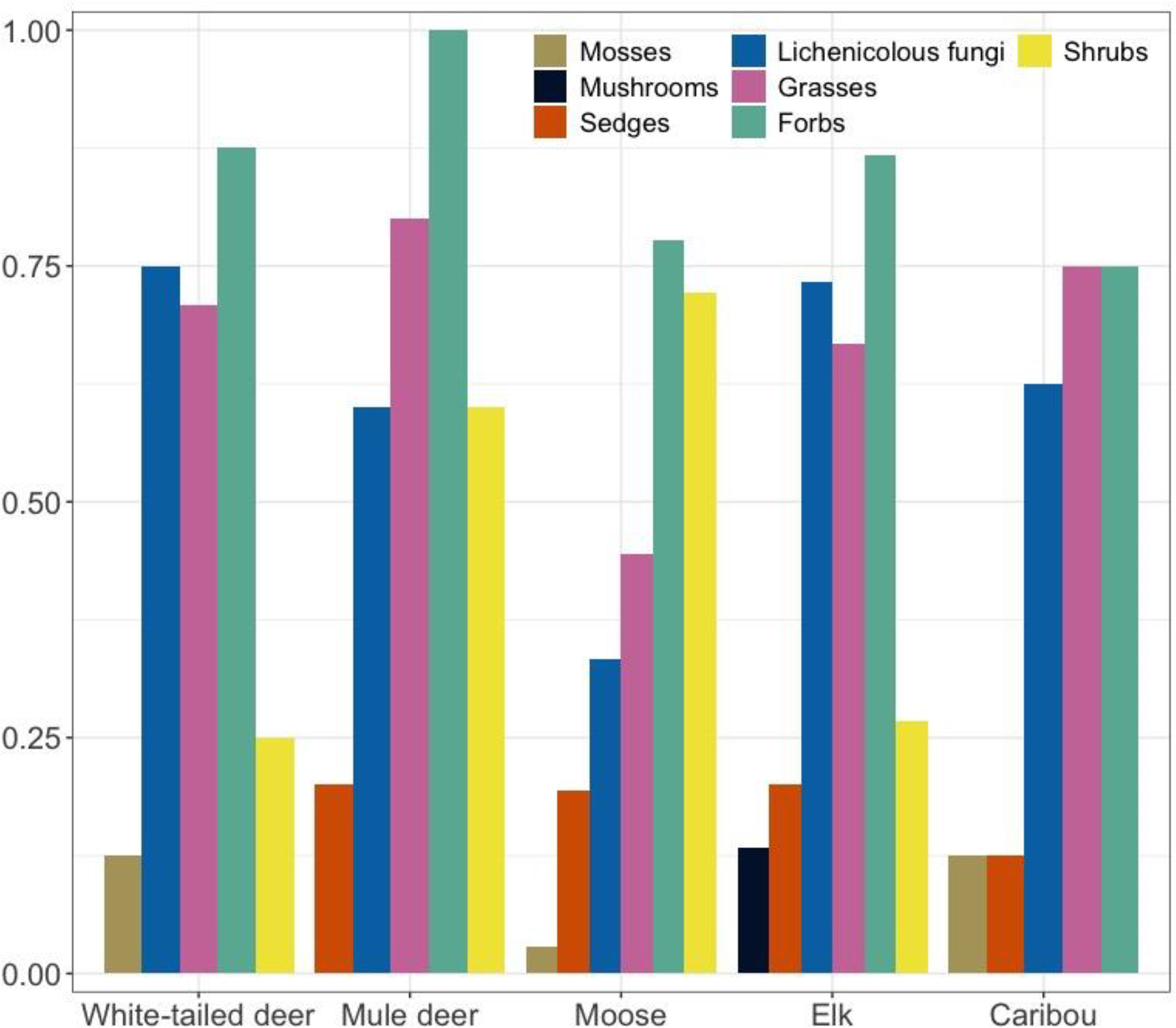
Lichenicolous fungi, grasses and forbs were identified in pellet samples from all five ungulates (Fig. 3). Shrubs/deciduous trees were identified in samples from all ungulates except caribou, and sedges were identified in samples from all ungulates except white-tailed deer. Mosses were found in pellet samples from white-tailed deer, moose, and caribou.
Fig. 3.
Across samples, the mean Horn-Morisita similarity index ranged from 0.4175 to 0.6402. Samples from the same ungulate were more similar to each other when compared to samples from other ungulates (Fig. 5). In general, moose samples had the lowest mean similarity with other ungulate samples and elk had the greatest mean similarity. These differences were typically not statistically significant, except when comparing moose with other ungulates (Fig. 6).
Diet abundance
The top five most abundant families of plants and fungi were similar for all five ungulates: Fabaceae (legumes), Lentibulariaceae (carnivorous plants, commonly butterworts), Nectriaceae (lichenicolous fungi), and Trichocomaceae (fungi associated with decaying material; Fig. 3; Supplemental Table 2). Acanthaceae (mint) was one of the most abundant families for moose. Considering diet items by forage group, forbs were the most abundant group for all five ungulates, comprising 53%–82% of the reads from plant DNA (Fig. 4). Shrubs/deciduous trees made up relatively little of the plant reads for deer and elk samples (2%–4%) and were substantially more abundant for moose (23% of plant reads). Lichenicolous fungi made up 23%–49% of the fungal DNA reads for each ungulate. Mosses, mushrooms, sedges, and grasses made up very little of each ungulate's diet.
Fig. 4.
Bray-Curtis similarity indices revealed that there was separation in diet among the five ungulate species (i.e., index <1.0) and that pellet samples from the same ungulate were more similar in diet to each other when compared to samples from other ungulates (Fig. 5). The exception to this pattern was white-tailed deer, where individual samples were more similar in diet to elk and caribou than to other white-tailed deer. However, the 95% confidence intervals overlapped for the species-species comparisons suggesting considerable imprecision in the raw data or results of the metabarcoding analysis. The average similarity index for each comparison ranged from 0.1951 to 0.4512 (Fig. 5). As with the Horn-Morisita index, moose had the lowest mean similarity index, but conversely to the Horn-Morisita results, caribou had the greatest mean similarity index when comparing abundance of each plant or fungal family. Using an ANOVA and post-hoc Dunnett Test, we found these differences to be statistically significant for moose compared to all other ungulates, for mule deer compared to white-tailed deer and moose, for elk compared to white-tailed deer, mule deer, and moose, and for caribou compared to elk, moose, and white-tailed deer (Fig. 6).
Fig. 5.
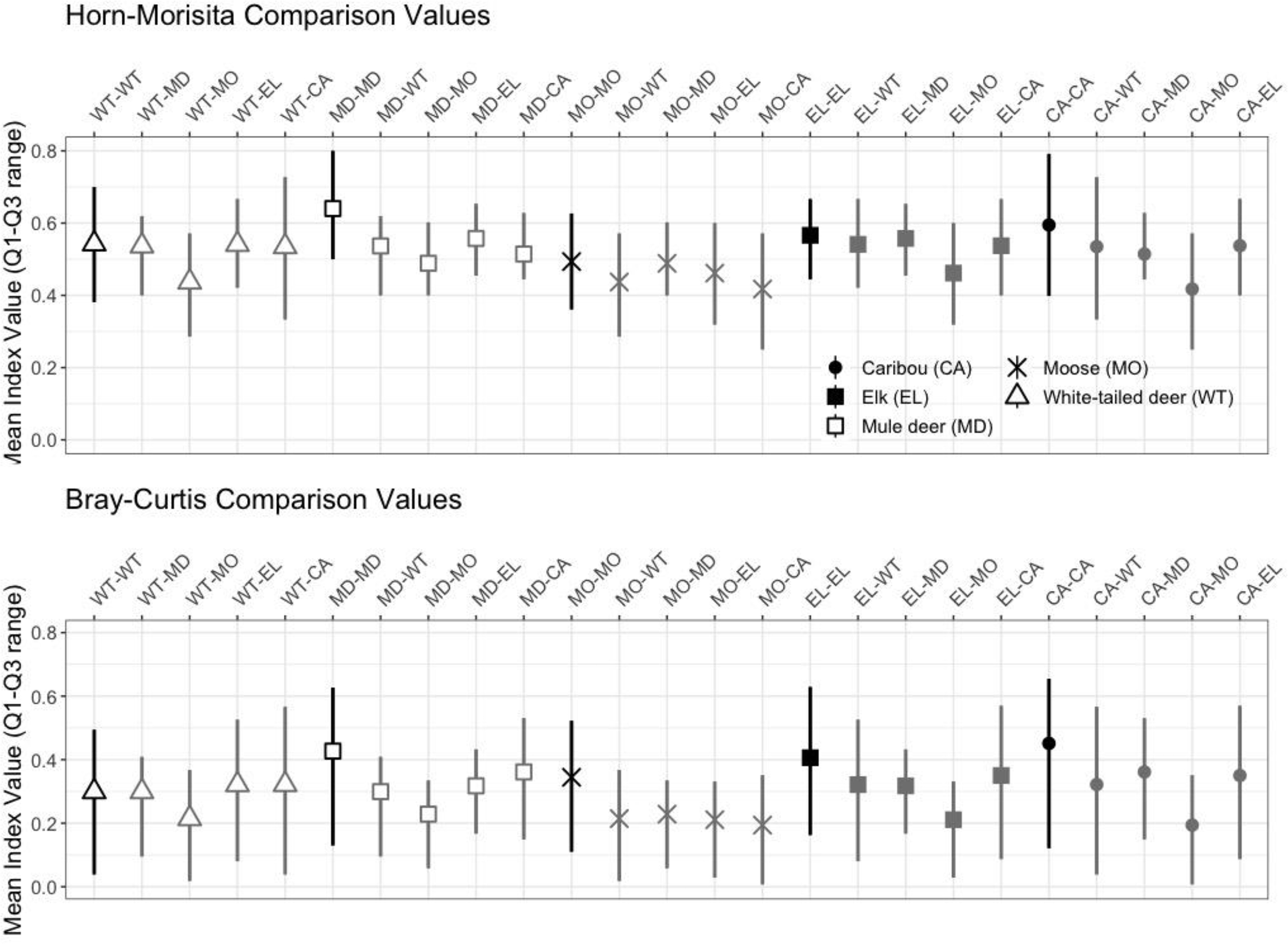
Fig. 6.
Discussion
This study provides the first diet comparison of the five forest-dwelling ungulates in west-central Alberta. We found that forbs were common in the diet of all five ungulates in this region during winter. The diet of the study ungulates had an overlap of 21%–30% when considering the abundance of plant and fungal families. However, this overlap was less than reported in several previously published studies (41%–95%: Hobbs et al. 1983; Leslie et al. 1984; Singer and Norland 1994; Kirchhoff and Larsen 1998; Torstenson et al. 2006), but similar to research from northwestern British Columbia and the Yukon (Hodder et al. 2013; Jung et al. 2015). For example, Hodder et al. (2013) reported that the diet of moose and elk overlapped by 11%, moose and mule deer by 24%, and elk and mule deer by 31%. In our study, moose had the most distinct diet (Horn-Morisita overlap 42%–49%, Bray-Curtis overlap 19%–22%), likely due to the abundance of Betula spp. that was relatively rare in the diets of the other four ungulate species.
Our results corroborate recent findings that caribou have a more varied diet than previously thought (Denryter et al. 2017; Mitchell et al. 2022; Webber et al. 2022). In particular, the diet of sampled caribou was not dominated by lichen during winter, in comparison to previous research in our study area (Thomas et al. 1996). We found that 75% of caribou samples contained grass and forb DNA, while 60% of the samples contained DNA associated with lichenicolous fungi. Unfortunately, the resolution of the analysis did not allow for an identification of specific genera that are known forage items of caribou (e.g., Cladina spp., Cladonia spp., Stereocaulon spp.; Thomas et al. 1996; Environment Canada 2014), so we could only identify the family of fungi that may be associated with those more specific forage species. In contrast to previous studies (summarized in Webber et al. 2022), the metabarcoding did not identify DNA of shrubs or deciduous trees in the caribou pellets that we collected. Also, we had a relatively small sample of pellets (N = 8), greatly limiting the generalizability of the findings to the population as well as woodland caribou, more generally.
Relative to the other four ungulates, moose had the greatest abundance of shrubs and deciduous trees, primarily from the Betula family, in their diet. In many regions, moose rely on conifer species such as fir during winter (Hodder et al. 2013). However, we found no conifer in any of the samples that we collected. It is possible that conifers were not a significant component of ungulate diets as the typical forage species (Abies spp., Taxus spp., Pseudotsuga mensiezii) were relatively uncommon or absent from our study area. Elk are often associated with grass (Holsworth 1960; Woods 1972; Salter and Hudson 1980; Churchill 1982; Kohl et al. 2012); however, we found deer, elk, and caribou had similar occurrence of grass species in their diet, and grass was only a small portion of the DNA reads for the individuals we sampled. We found that the diet of white-tailed deer and mule deer was varied, which was expected for these species, but the sample size was extremely small for mule deer (Franzmann and Schwartz 1997; Hewitt 2011).
Limitations
The results of this study were informative but must be considered in the context of the limitations of sampling and DNA metabarcoding of fecal samples. Although we have no measure of absolute diet for the species or study area that we sampled, there is the possibility of misclassification of taxonomic family. Indeed, a number of the results were surprising. For example, there was no evidence of conifer trees (Pinaceae) in any of the pellet samples. Previous diet analyses from western Canada reported that moose and deer consumed Abies spp., Pinus spp., and Douglas-fir during winter (Hodder et al. 2013; Rea 2014; Rea et al. 2017; Koetke et al. 2023). Also, the laboratory analysis revealed some plants that were likely very rare across the study area during winter. For example, Lentibulariaceae, including butterworts and bladderworts, are small flowering plants typically found in wet aquatic habitat. We must assume that the shoots of those plants would be unavailable as forage during winter.
Sampling of fecal pellets occurred soon after snowfall. This resulted in uneven sample sizes and non-random distribution of the sample locations. Collection sites may be biased towards proximity to roads, which can influence the availability of plant species and ultimately diet (Roever et al. 2008). The small number of samples (N = 88 for five species) resulted in limited generalizability of the findings. Also, the sample was unbalanced, with relatively few samples for caribou (N = 8) and mule deer (N = 5). Finally, animal diets vary spatially and over time (e.g., Bojarska and Selva 2012; DeBano et al. 2016; Koetke et al. 2023). Future work in our study area would benefit from greater and seasonal sampling that more fully represents spatial variation as well as within and among season differences in diets.
Pellet samples for caribou were provided from previous projects and had been frozen for five years prior to analysis. Long-term storage could have influenced the viability of the DNA with a differential effect of degradation among plant and fungal types (Nsubuga et al. 2004). However, previous research has shown limited differences in DNA extracted from samples frozen for multiple years (Gavriliuc et al. 2021). Our results were based on fungal DNA present in the samples, which was then inferred to be lichenicolous based on fungal associations (Mitchell et al. 2022). We assumed fungal DNA was consumed purposefully by the ungulate. However, it is possible that fungi were not targeted forage items but were consumed incidentally (e.g., endophytic) or fungal spores settled out on pellets after defecation. While DNA metabarcoding is expected to provide higher taxonomic resolution than methods such as microhistology (Port et al. 2016), we found only 30% of the fungal reads and 60% of the plant reads identified to the genus level.
Key messages
Forage and forage subsidies have garnered attention for their role in apparent competition between caribou and white-tailed deer (Fisher and Burton 2021; Fuller et al. 2023). That interaction is thought to be facilitated by less severe winters and an increase in the availability of early-seral plants resulting from human-caused land clearing (e.g., forestry, oil and gas extraction). Despite those concerns, there has been no research into the diet of white-tailed deer in this region. Our study provides some insight into the diet of white-tailed deer in west-central Alberta, and the first comparative analysis of the diet of caribou and four possible apparent competitors. We found that vascular plants, specifically forbs, were a substantial component of each ungulate’s diet. This commonality in diet may influence the overlap between caribou and the other ungulate species in this system.
Competitive exclusion occurs when two species occupy the same niche, resulting in one species being excluded (Armstrong and McGehee 1980). Our results suggest that there is a relatively low risk of these five ungulates occupying the same forage niche, but there may be competition for specific food items. Caribou rely predominately on lichen for winter forage (Thomas et al. 1996), and we found lichenicolous fungi in >30% of the samples for each ungulate. Specifically, Nectriaceae made up >25% of the fungal DNA reads. This corroborates other evidence that many ungulates use lichen for winter forage (Thomas 1990; Latham and Boutin 2008; Hodder et al. 2013). Our results provide some evidence of possible exploitative competition between caribou and other ungulates in west-central Alberta. However, we have no understanding of the total consumption of shared plants or lichens among the five ungulates that we studied. Similar family groups within pellet samples does not mean that forage is limiting or that niche overlap has a direct fitness effect for caribou, deer, elk, or moose. In theory, these ungulates could have an identical proportion of each family group within their pellets, but diet could be differentiated at the species level.
We reported some ecologically plausible patterns in the diet of the five species in this study, but indeterminate results suggested that there was still some need to evaluate the efficacy and accuracy of DNA metabarcoding as a method for quantifying diet represented in fecal pellets. For our small sample, we achieved poor taxonomic resolution, relative to other studies and methods. Also, we suspected some misclassification of plant and fungal families. Nonetheless, this method is relatively efficient and could provide a foundation to explore how the concurrent effects of climate and landscape change, among other factors, may influence the nutritional ecology of sympatric Cervidae.
Acknowledgements
We thank the staff at fRI Research for collecting and storing caribou pellets from 2016, as well as staff from the Government of Alberta who facilitated pellet collection in 2022. The Canadian Center for DNA Barcoding and Wildlife Genetics International conducted the laboratory analyses.
References
Alberta Agriculture and Forestry. 2017. Sustainable forest management: 2016 facts & statistics, annual allowable cut. Available from https://open.alberta.ca/publications/2368-4895.
Ando H., Mukai H., Komura T., Dewi T., Ando M., Isagi Y. 2020. Methodological trends and perspectives of animal dietary studies by noninvasive fecal DNA metabarcoding. Environmental DNA, 2(4): 391–406.
Armstrong R.A., McGehee R. 1980. Competitive exclusion. The American Naturalist, 115(2): 151–170.
Bach B.H., Quigley A.B., Gaynor K.M., McInturff A., Charles K.L., Dorcy J., Brashares J.S. 2022. Identifying individual ungulates from fecal DNA: a comparison of field collection methods to maximize efficiency, ease, and success. Mammalian Biology, 102(3): 863–874.
Bischof R., Loe L.E., Meisingset E.L., Zimmermann B., Van Moorter B., Mysterud A. 2012. A migratory northern ungulate in the pursuit of spring: jumping or surfing the green wave? The American Naturalist, 180(4): 407–424.
Boan J.J., McLaren B.E., Malcolm J.R. 2011. Influence of post-harvest silviculture on understory vegetation: implications for forage in a multi-ungulate system. Forest Ecology and Management, 262(9): 1704–1712.
Bojarska K., Selva N. 2012. Spatial patterns in brown bear Ursus arctos diet: the role of geographical and environmental factors. Mammal Review, 42(2): 120–143.
Bonin M., Dussault C., Taillon J., Lecomte N., Côté S. 2020. Combining stable isotopes, morphological, and molecular analyses to reconstruct the diet of free-ranging consumers. Ecology and Evolution, 10(13): 6664–6676.
Bradshaw R.H.W., Hannon G.E., Lister A.M. 2003. A long-term perspective on ungulate–vegetation interactions. Forest Ecology and Management, 181(1): 267–280.
Bray J.R., Curtis J.T. 1957. An ordination of the upland forest communities of southern Wisconsin. Ecological Monographs, 27(4): 325–349.
Brecka A.F.J., Shahi C., Chen H.Y.H. 2018. Climate change impacts on boreal forest timber supply. Forest Policy and Economics, 92(C): 11–21.
Brodo I.M., Sharnoff S.D., Sharnoff S. 2001. Lichens of North America. Blue Jay, 60(1).
Churchill B.P. 1982. Winter habitat selection and use of clearcuts by elk in the White River Drainage of southeastern British Columbia. M.Sc. thesis, University of Victoria, Victoria, British Columbia.
COSEWIC. 2014. COSEWIC assessment and status report on the Caribou Rangifer tarandus, Northern Mountain population, Central Mountain population and Southern Mountain population in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. xxii + 113pp.
de Sousa L.L., Silva S.M., Xavier R. 2019. DNA metabarcoding in diet studies: Unveiling ecological aspects in aquatic and terrestrial ecosystems. Environmental DNA 1(3): 199–214.
Deagle B.E., Thomas A.C., McInnes J.C., Clarke L.J., Vesterinen E.J., Clare E.L., et al. 2019. Counting with DNA in metabarcoding studies: how should we convert sequence reads to dietary data? Molecular Ecology, 28(2): 391–406.
DeBano S.J., Roof S.M., Rowland M.M., Smith L.A. 2016. Diet overlap of mammalian herbivores and native bees: implications for managing co-occurring grazers and pollinators. Natural Areas Journal, 36(4): 458–477.
DeCesare N.J., Hebblewhite M., Robinson H.S., Musiani M. 2010. Endangered, apparently: the role of apparent competition in endangered species conservation. Animal Conservation, 13(4): 353–362.
Denryter K.A., Cook R.C., Cook J.G., Parker K.L. 2017. Straight from the caribou's (Rangifer tarandus) mouth: detailed observations of tame caribou reveal new insights into summer–autumn diets. Canadian Journal of Zoology, 95(2): 81–94.
Dyer S.J., O'Neil J.P., Wasel S.M., Boutin S. 2001. Avoidance of industrial development by woodland caribou. Journal of Wildlife Management, 65(3): 531–542.
Edenius L., Månsson J., Hjortstråle T., Roberge J.-M., Ericsson G. 2015. Browsing and damage inflicted by moose in young Scots pine stands subjected to high-stump precommercial thinning. Scandinavian Journal of Forest Research, 30(5): 1–6.
Edenius L., Roberge J.-M., Månsson J., Ericsson G. 2014. Ungulate-adapted forest management: effects of slash treatment at harvest on forage availability and use. European Journal of Forest Research, 133(1): 191–198.
Environment Canada. 2014. Recovery Strategy for the Woodland Caribou, Southern Mountain Population (Rangifer Tarandus Caribou) in Canada. Species at Risk Act Recovery Strategy Series. Environment Canada, Ottawa. viii + 103pp.
Finnegan L., Pigeon K.E., MacNearney D. 2019. Predicting patterns of vegetation recovery on seismic lines: informing restoration based on understory species composition and growth. Forest Ecology and Management, 446(1): 175–192.
Fisher J.T., Burton A.C. 2021. Spatial structure of reproductive success infers mechanisms of ungulate invasion in Nearctic boreal landscapes. Ecology and Evolution, 11(1): 900–911.
Fisher J.T., Burton A.C., Nolan L., Roy L. 2020. Influences of landscape change and winter severity on invasive ungulate persistence in the Nearctic boreal forest. Scientific Reports, 10(1): 8742.
Franzmann A.W., Schwartz C.C. (eds). 1997. Ecology and management of the North American moose. 1st ed. Smithsonian Institution Press, Washington, D.C.
Fuller H.W., Frey S., Fisher J.T. 2023. Integration of aerial surveys and resource selection analysis indicates human land use supports boreal deer expansion. Ecological Applications, 33(1): e2722.
Gavriliuc S., Stothart M.R., Henry A., Poissant J. 2021. Long-term storage of feces at −80 °C versus −20 °C is negligible for 16S rRNA amplicon profiling of the equine bacterial microbiome. PeerJ, 9(2): e10837.
Government of Alberta. 2022. Alberta Conservation Information Management System (ACIMS). Available from https://www.albertaparks.ca/albertaparksca/management-land-use/alberta-conservation-information-management-system-acims/.
Haeussler S., Bedford L., Leduc A., Bergeron Y., Kranabetter J. 2002. Silvicultural disturbance severity and plant communities of the southern Canadian boreal forest. Silva Fennica, 36(1): 307–327.
Hansen M.C., Potapov P.V., Moore R., Hancher M., Turubanova S.A., Tyukavina A., et al. 2013. High-resolution global maps of 21st-century forest cover change. Science, 342(6160): 850–853.
Hebblewhite M., Merrill E., McDermid G. 2008. A multi-scale test of the forage maturation hypothesis in a partially migratory ungulate population. Ecological Monographs, 78(2): 141–166.
Hewitt D.G. editor. 2011. Biology and management of White-tailed deer. CRC Press, Boca Raton, Florida.
Hobbs N.T., Baker D.L., Gill R.B. 1983. Comparative nutritional ecology of montane ungulates during winter. The Journal of Wildlife Management, 47(1): 1–16.
Hodder D.P., Rea R.V., Crowley S.M. 2013. Diet content and overlap of sympatric mule deer (Odocoileus hemionus), Moose (Alces alces), and elk (Cervus elaphus) during a deep snow winter in north- central British Columbia, Canada. Canadian Wildlife Biologist and Management 2(2): 43–50.
Holsworth W.N. 1960. Interactions between moose, elk and buffalo in Elk Island National Park, Alberta. M.Sc. thesis, University of British Columbia, Vancouver, British Columbia.
Horn H.S. 1966. Measurement of ‘overlap’ in comparative ecological studies. The American Naturalist, 100(914): 419–424.
Ivanova N.V., Dewaard J.R., Hebert P.D.N. 2006. An inexpensive, automation-friendly protocol for recovering high-quality DNA. Molecular Ecology Notes, 6(4): 998–1002.
Jackson N.J., Stewart K.M., Wisdom M.J., Clark D.A., Rowland M.M. 2021. Demographic performance of a large herbivore: effects of winter nutrition and weather. Ecosphere, 12(1): e03328.
Johnson C.J., Ehlers L.P.W., Seip D.R. 2015. Witnessing extinction—cumulative impacts across landscapes and the future loss of an evolutionarily significant unit of woodland caribou in Canada. Biological Conservation, 186(1): 176–186.
Johnson C.J., Parker K.L., Heard D.C. 2001. Feeding site selection by woodland caribou in north-central British Columbia. Rangifer, 20(5): 158–172.
Jung T.S., Stotyn S.A., Czetwertynski S.M. 2015. Dietary overlap and potential competition in a dynamic ungulate community in Northwestern Canada. The Journal of Wildlife Management, 79(8): 1277–1285.
King S.R.B., Schoenecker K.A. 2019. Comparison of methods to examine diet of feral horses from noninvasively collected fecal samples. Rangeland Ecology & Management, 72(4): 661–666.
Kirchhoff M.D., Larsen D.N. 1998. Dietary overlap between native sitka black-tailed deer and introduced elk in Southeast Alaska. The Journal of Wildlife Management, 62(1): 236–242.
Koetke L.J., Hodder D.P., Rea R.V., Johnson C.J., Marshall S. 2023. Landscape disturbance alters the composition and diversity of the diet of moose, a generalist herbivore. Forest Ecology and Management, 530(1): 120760.
Kohl M.T., Hebblewhite M., Cleveland S.M., Callaway R.M. 2012. Forage value of invasive species to the diet of rocky mountain elk. Rangelands, 34(2): 24–28.
Latham A.D.M., Boutin S. 2008. Evidence of arboreal lichen use in peatlands by white-tailed deer, Odocoileus virginianus, in northeastern Alberta. The Canadian Field-Naturalist, 122(3): 230–233.
Lawrey J.D., Diederich P. 2018. Lichenicolous fungi: Worldwide checklist, including isolated cultures and sequences. http://www.lichenicolous.net/.
Leslie D.M., Starkey E.E., Vavra M. 1984. Elk and deer diets in old-growth forests in western Washington. The Journal of Wildlife Management, 48(3): 762–775.
Littleford-Colquhoun B.L., Freeman P.T., Sackett V.I., Tulloss C.V., McGarvey L.M., Geremia C., Kartzinel T.R. 2022. The precautionary principle and dietary DNA metabarcoding: commonly used abundance thresholds change ecological interpretation. Molecular Ecology, 31(6): 1615–1626.
Mitchell G., Wilson P.J., Manseau M., Redquest B., Patterson B.R., Rutledge L.Y. 2022. DNA metabarcoding of faecal pellets reveals high consumption of yew (Taxus spp.) by caribou (Rangifer tarandus) in a lichen-poor environment. FACETS, 7(1): 701–717.
Monteith K.L., Stephenson T.R., Bleich V.C., Conner M.M., Pierce B.M., Bowyer R.T. 2013. Risk-sensitive allocation in seasonal dynamics of fat and protein reserves in a long-lived mammal. Journal of Animal Ecology, 82(2): 377–388.
Moran A.J., Prosser S.W.J., Moran J.A. 2019. DNA metabarcoding allows non-invasive identification of arthropod prey provisioned to nestling rufous hummingbirds (Selasphorus rufus). PeerJ, (7): e6596.
Natural Regions Committee 2006. Natural Regions and Subregions of Alberta. Compiled by DJ Downing and WW Pettapiece. Government of Alberta. Pub. No. T/852.
Nichols R.V., Cromsigt J., Spong G. 2015. Using eDNA to experimentally test ungulate browsing preferences. SpringerPlus, 4(1): 489.
Nowlin R.A. 1978. Habitat selection and food habits of moose in northeastern Alberta. Alces: A Journal Devoted to the Biology and Management of Moose, 14(1): 178–193.
Nsubuga A.M., Robbins M.M., Roeder A.D., Morin P.A., Boesch C., Vigilant L. 2004. Factors affecting the amount of genomic DNA extracted from ape faeces and the identification of an improved sample storage method. Molecular Ecology, 13(7): 2089–2094.
Oksanen J., Simpson G., Blanchet F., Kindt R., Legendre P., Minchin P., et al. 2022. vegan: Community Ecology Package. R package version 2.6-4. https://CRAN.R-project.org/package=vegan.
Park H., Jeong S., Peñuelas J. 2020. Accelerated rate of vegetation green-up related to warming at northern high latitudes. Global Change Biology, 26(11): 6190–6202.
Paula D.P., Barros S.K.A., Pitta R.M., Barreto M.R., Togawa R.C., Andow D.A. 2022. Metabarcoding versus mapping unassembled shotgun reads for identification of prey consumed by arthropod epigeal predators. GigaScience, 11(1): giac020.
Pickell P.D., Andison D.W., Coops N.C., Gergel S.E., Marshall P.L. 2015. The spatial patterns of anthropogenic disturbance in the western Canadian boreal forest following oil and gas development. Canadian Journal of Forest Research, 45(6): 732–743.
Pimm S.L., Diamond J., Reed T.M., Russell G.J., Verner J. 1993. Times to extinction for small populations of large birds. Proceedings of the National Academy of Sciences, 90(22):10871–10875.
Pompanon F., Deagle B.E., Symondson W.O.C., Brown D.S., Jarman S.N., Taberlet P. 2012. Who is eating what: diet assessment using next generation sequencing. Molecular Ecology, 21(8): 1931–1950.
Port J.A., O'Donnell J.L., Romero-Maraccini O.C., Leary P.R., Litvin S.Y., Nickols K.J., et al. 2016. Assessing vertebrate biodiversity in a kelp forest ecosystem using environmental DNA. Molecular Ecology, 25(2): 527–541.
Post E., Steinman B.A., Mann M.E. 2018. Acceleration of phenological advance and warming with latitude over the past century. Scientific Reports, 8(1): 3927.
Proffitt K.M., Hebblewhite M., Peters W., Hupp N., Shamhart J. 2016. Linking landscape-scale differences in forage to ungulate nutritional ecology. Ecological Applications, 26(7): 2156–2174.
QGIS Development Team. 2021. QGIS geographic information system. QGIS Association. https://www.qgis.org.
R Core Team. 2022. R: A language and environment for statistical computing. R Foundation for Statistical Computing. Vienna, Austria. https://www.R-project.org/.
Raubenheimer D., Simpson S.J., Mayntz D. 2009. Nutrition, ecology and nutritional ecology: toward an integrated framework. Functional Ecology, 23(1): 4–16.
Rea R.V. 2014. A preliminary analysis of moose (Alces alces) winter diets in the Aleza Lake Research Forest in north-central British Columbia. Wildlife Afield, 11(1): 50–53.
Rea R.V., Hjeljord O., Langen P. 2017. Conifer diet choices made by mule deer (Odocoileus hemionus) of north central British Columbia during a cafeteria-style feeding trial. Northwest Science, 91(1): 90–99.
Renecker L.A., Hudson R.J. 1986. Seasonal foraging rates of free-ranging moose. The Journal of Wildlife Management, 50(1): 143–147.
Renecker L.A., Hudson R.J. 1988. Seasonal quality of forages used by moose in the aspen-dominated boreal forest, central Alberta. Holarctic Ecology, 11(2): 111–118.
Roever C.L., Boyce M.S., Stenhouse G.B. 2008. Grizzly bears and forestry II: Grizzly bear habitat selection and conflicts with road placement. Forest Ecology and Management, 256(6): 1262–1269.
Sala O.E., Stuart Chapin F., Armesto J.J., Berlow E., Bloomfield J., Dirzo R., et al. 2000. Global biodiversity scenarios for the year 2100. Science, 287(5459): 1770–1774.
Salter R.E., Hudson R.J. 1980. Range relationships of feral horses with wild ungulates and cattle in western Alberta. Journal of Range Management, 33(4): 266.
Singer F.J., Norland J.E. 1994. Niche relationships within a guild of ungulate species in Yellowstone National Park, Wyoming, following release from artificial controls. Canadian Journal of Zoology, 72(8): 1383–1394.
Soja A.J., Tchebakova N.M., French N.H.F., Flannigan M.D., Shugart H.H., Stocks B.J., et al. 2007. Climate-induced boreal forest change: predictions versus current observations. Global and Planetary Change, 56(3–4): 274–296.
Strong W.L., Gates C.C. 2006. Herbicide-induced changes to ungulate forage habitat in western Alberta, Canada. Forest Ecology and Management, 222(1–3): 469–475.
Sulla-Menashe D., Woodcock C.E., Friedl M.A. 2018. Canadian boreal forest greening and browning trends: an analysis of biogeographic patterns and the relative roles of disturbance versus climate drivers. Environmental Research Letters, 13(1): 014007.
Tattersall E., Pigeon K., MacNearney D., Finnegan L. 2023. Walking the line: investigating biophysical characteristics related to wildlife use of linear features. Ecological Solutions and Evidence, 4(1): e12219.
Thomas D.C. 1990. Moose diet and use of successional forests in the Canadian taiga. Alces: A Journal Devoted to the Biology and Management of Moose, 26(1): 24–29.
Thomas D.C., Edmonds E.J., Brown W.K. 1996. The diet of woodland caribou populations in west-central Alberta. Rangifer, 16(4): 337.
Thompson I.D., Wiebe P.A., Mallon E., Rodgers A.R., Fryxell J.M., Baker J.A., et al. 2015. Factors influencing the seasonal diet selection by woodland caribou (Rangifer tarandus tarandus) in boreal forests in Ontario. Canadian Journal of Zoology, 93(2): 87–98.
Torstenson W.L.F., Mosley J.C., Brewer T.K., Tess M.W., Knight J.E. 2006. Elk, mule deer, and cattle foraging relationships on foothill and mountain rangeland. Rangeland Ecology & Management, 59(1): 80–87.
Webber Q.M.R., Ferraro K.M., Hendrix J.G., Vander Wal E. 2022. What do caribou eat? A review of the literature on caribou diet. Canadian Journal of Zoology, 100(3): 197–207.
Weisberg P.J., Bugmann H. 2003. Forest dynamics and ungulate herbivory: from leaf to landscape. Forest Ecology and Management, 181(1–2): 1–12.
Willams W., McLean A., Tucker R., Ritcey R. 1980. Deer and cattle diets on summer range in British Columbia. Journal of Range Management, 33(1): 55–59.
Woods J.G. 1972. Ecology of a partially migratory elk population. PhD dissertation, University of British Columbia, Vancouver, British Columbia.
Supplementary material
Supplementary Material 1 (DOCX / 46 KB).
- Download
- 45.38 KB
Information & Authors
Information
Published In

FACETS
Volume 10 • 2025
Pages: 1 - 13
Editor: Allyson Kate Menzies
History
Received: 31 October 2023
Accepted: 3 October 2024
Version of record online: 31 January 2025
Copyright
© 2025 The Authors. This work is licensed under a Creative Commons Attribution 4.0 International License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Data Availability Statement
Data generated or analyzed during this study are available from the corresponding author upon reasonable request.
Key Words
Sections
Subjects
Authors
Author Contributions
Conceptualization: SS, CJJ, RVR
Data curation: SS
Formal analysis: SS, CJJ, LF
Funding acquisition: CJJ, LF
Investigation: SS
Methodology: SS, CJJ
Project administration: CJJ
Resources: LF
Supervision: CJJ, LF
Validation: SS, CJJ
Visualization: SS
Writing – original draft: SS
Writing – review & editing: CJJ, LF, RVR
Competing Interests
The authors have no competing interests to declare.
Funding Information
fRI Research
Mitacs: IT26597
Sustainable Forestry Initiative: SFI, 2013-03
Alberta Upstream Petroleum Research Fund: 15-ERPC-01
Weyerhaeuser Company Ltd.
Forest Resource Improvement Association of Alberta: FRIP, OF-12-006
Funding for this project was provided by a Mitacs Accelerate Grant (IT26597), Natural Sciences and Engineering Research Council of Canada, fRI Research, partners of the fRI Research Caribou Program, and the Forest Resource Improvement Association of Alberta [FRIAA] (FOOMOD-01-054). Caribou fecal collection and laboratory analysis was supported by the Sustainable Forestry Initiative (SFI 2013-03), FRIAA (FRIP OF-12-006), the Alberta Upstream Petroleum Research Fund (15-ERPC-01), and Weyerhaeuser Company Ltd.
Metrics & Citations
Metrics
Other Metrics
Citations
Cite As
Suzanne Stevenson, Chris J. Johnson, Laura Finnegan, and Roy V. Rea. 2025. Winter diet of five sympatric ungulates in west-central Alberta, Canada—inference from DNA metabarcoding of fecal pellets. FACETS.
10: 1-13.
https://doi.org/10.1139/facets-2023-0195
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
There are no citations for this item