Transport does not influence maximum metabolic rate or thermal tolerance of endangered redside dace (Clinostomus elongatus)
Abstract
Reintroduction is an important tool in the conservation and recovery of aquatic species at risk. However, components of the reintroduction process such as transportation have the potential to induce physiological stress and the extent to which preparatory techniques can mitigate this stress is poorly understood in small-bodied fishes. To address this concern, we studied the effect of transport on two fitness-related performance measures: maximum metabolic rate and thermal tolerance in redside dace (Clinostomus elongatus), an imperilled small-bodied stream fish native to eastern North America. Prior to transportation, we manipulated the body condition of redside dace over a 12-week period, by providing either low (1% of their total body mass) or high (2% of their total body mass) rations. The goal of this manipulation was to influence body condition, as higher body condition can enhance physiological performance. Subsequently, redside dace were transported for varying durations: 0, 3, and 6 h. Following transportation, we measured maximum metabolic rate (µmol/h) and thermal tolerance (CTmax, °C). Our results indicate that neither transport nor body condition had a significant effect on maximum metabolic rate or thermal tolerance (CTmax). These findings provide preliminary evidence that redside dace can physiologically tolerate transport based on the endpoints measured and this information may possibly be extended to other small-bodied fish, for which information is lacking.
Introduction
Reintroduction is an important tool for the conservation and recovery of aquatic species at risk (Seddon et al. 2007; Cochran-Biederman et al. 2015; Lamothe et al. 2019; Drake et al. 2021). Reintroduction success ultimately hinges on the long-term survival and reproduction of released individuals. One of the most critical periods for reintroduced animals occurs during the first hours after release, when rapid acclimatization to a novel environment is required (Näslund 2021). This period may be especially difficult because of the repeated stress imparted by reintroduction procedures (Dickens et al. 2010). For fishes in particular, transportation can be particularly challenging, causing a suite of physiological changes due to handling, air exposure, confinement, motion during transport, changes in water quality parameters, and high conspecific densities (Teixeira et al. 2007; Sampaio and Freire 2016). Accordingly, the potential for components of the reintroduction process to induce behavioral and physiological stress must be carefully considered, and strategies must be implemented to minimize their negative consequences.
One promising strategy to alleviate some of the negative effects of transport is the provisioning of extra food in the weeks prior to transport to bolster condition, which may help increase physiological resilience. When fish encounter acute stressors (e.g., those associated with transport), the activation of the HPI (hypothalamic-pituitary-interrenal) axis triggers the production and circulation of glucocorticoids, energy-mobilizing hormones that shift energy resources away from regular physiological processes, redirecting them toward restoring homeostasis (Barton et al. 2002). This stress-induced physiological response is energetically costly (Barton 2002); however, the extent of these negative costs may depend on the nutritional state of fish. For instance, food availability may improve physiological performance measures such as maximum metabolic rate (MMR) and thermal tolerance, two fitness-related measures of physiological resilience thought to be influenced by body condition (e.g., Pörtner and Peck 2010; Norin and Clark 2016; Turko et al. 2020). Accordingly, during the captive rearing stage of the reintroduction process, there is a unique opportunity to increase body condition through supplemental feeding or other dietary adjustments. Although much research has explored supplementation of specific dietary components such as carotenoids, probiotics, and glucan (see review in Vanderzwalmen et al. 2019), there has been limited focus on overall condition factor. While energy storage and food availability can reduce energy demands during stressful events (McEwen and Wingfield 2003), a better understanding of whether nutrition and dietary strategies can play a role in the success of reintroduction efforts is warranted.
Measuring ecologically relevant indicators that reflect the connection between the stress response and fish performance following transport is important to improve the success of reintroductions. MMR (and subsequent recovery) is an ecologically important indicator of physiological resilience, representing the upper limit at which oxygen can be delivered from the environment to the mitochondria within an organism’s tissues (Norin and Clark 2016). At a minimum, organisms must allocate energy toward the maintenance of essential homeostatic mechanisms to sustain life—beyond this, any surplus can be allocated to other functions, albeit within the confines defined by MMR (Fry 1971). In essence, MMR sets an upper boundary on an organism’s ability to perform oxygen-consuming physiological activities (Killen et al. 2017). MMR has gained considerable attention across a variety of contexts in the lab and field (Norin and Clark 2016), and could prove particularly valuable for assessments of imperilled species, which are likely to be vulnerable to the stressors associated with transportation (Seddon et al. 2012). When organisms are released into unfamiliar environments, they must immediately confront two physiologically demanding challenges: the need to disperse in search of food resources and evading native predators (Näslund 2021). If MMR is compromised as a result of transport stress, it could subsequently heighten the vulnerability of reintroduced fish. However, whether transport can reduce MMR, potentially increasing the underlying physiological vulnerability during reintroduction, remains unclear.
Along with finding food and evading predators, reintroduced fish must also cope with novel stressors in their unfamiliar environment, such as temperature fluctuations (Sébastien et al. 2021). Critical thermal maximum (CTmax) is a performance metric reflecting physiological resilience that is used to assess the thermal tolerance of fishes. CTmax is the temperature at which an animal loses equilibrium, signifying an “ecological death” under natural conditions (Lutterschmidt and Hutchison 1997; Schulte et al. 2011). Stress can adversely affect CTmax, resulting in a decreased ability to tolerate temperature extremes or narrow the range of temperatures for fish to function optimally (e.g., Monirian et al. 2010; LeBlanc et al. 2011). For example, in a study on Threadfin shad (Dorosoma petenense), handling stress was found to significantly affect both survival and the ability to tolerate varying temperatures, resulting in higher mortality rates and lower CTmax compared to non-handled control fish (Monirian et al. 2010). Thus, as the success of reintroduction efforts often hinges on the mitigation of stress-induced mortality and ensuring a species’ capacity to adapt to a novel environment, CTmax, like MMR, may be an important metric to consider.
The redside dace (Clinostomus elongatus) is a small minnow species found in streams throughout northeastern North America (Redside Dace Recovery Team 2010; COSEWIC 2017). Populations of these minnows have significantly declined, leading to their imperilled status across much of their United States range (reviewed in Serrao et al. 2018) and their designation as Endangered in Canada (Redside Dace Recovery Team 2010; COSEWIC 2017). In the wild, redside dace face many threats, including thermal stress resulting from the impact of urbanization on nearby waterbodies, compounded by the effects of climate change (Parker et al. 1988; Redside Dace Recovery Team 2010; COSEWIC 2017; Turko et al. 2020). In response to this conservation concern, captive breeding programs have been initiated in Ontario, Canada to address the decline and offer potential solutions, such as supplementing existing wild populations and providing individuals for future reintroductions. As a result, the transportation of individuals from captivity to the wild and vice versa will require appropriate techniques and strategies to ensure survival.
As reintroduction efforts for redside dace will require transport events, and neither the physiological impact of transportation stress nor potential stress-reduction strategies have been evaluated, the objective of this study is to explore the potential impacts of transport on ecologically relevant physiological traits that may be closely linked to fitness immediately after reintroduction. We hypothesize that transport will lead to changes in metabolic rate. We predict that transported redside dace will exhibit both a lower MMR and an extended period of recovery, but that high body condition fish will have higher MMR than low body condition fish, regardless of transportation. We also hypothesize that transport will reduce thermal tolerance (CTmax). We predict that as transport time increases, CTmax will decrease, but that high body condition fish will have higher CTmax than low body condition fish. Taken together, this study therefore aims to investigate the effect of a management-relevant transport event on redside dace using multiple ecologically meaningful indicators of stress, with potential future applications for species-specific reintroduction planning. This research further investigates whether employing preparatory techniques, namely, enhanced body condition through supplemental feeding, can mitigate some of the potentially negative physiological consequences of transport. More broadly, this work adds to the paucity of literature assessing transport stress in imperilled, small-bodied fishes in comparison to species with economic importance for stocking and aquaculture.
Materials and methods
Fish collection and husbandry
Between 2018 and 2019, redside dace were collected from the Kokosing River in Morrow County, Ohio, USA (coordinates: 40.545909, −82.654234) using standard seining techniques. Fish were then transported to the University of Windsor’s Freshwater Restoration Ecology Centre (FREC) in Lasalle, Ontario, Canada (Turko et al. 2020). Fish collection permits were issued by the Ohio Department of Natural Resources Division of Wildlife. Upon arrival to FREC, fish were housed in four fiberglass tanks (800 L, 914 mm deep × 1549 mm diameter) for approximately three and a half years prior to experimentation. The tanks were part of a recirculating system where approximately 10% of the water was exchanged daily. Water temperature ranged between 12 and 18 °C and pH ranged between 7.1 and 7.8. A light cycle of 12 h light: 12 h dark (with lights on at 0600 h) was maintained by overhead fluorescent lights. Two of the tanks included a loose layer of multicolored pebble gravel (3/8 in.; brown, gray, white) covering roughly 80% of the base of the tank (Wentworth 1922). Each enriched tank also contained vertical floating plastic plants (∼30 cm tall). The remaining two tanks had a barren environment absent of substrate and plants. All fish were fed commercial fish flakes 3–4 times per week (∼ 1%–2% of total body mass). All animals were cared for in accordance with the Guide to the Care and Use of Experimental Animals (available from the Canadian Council on Animal Care website: www.ccac.ca), and the use of animals was reviewed and approved by the Animal Care Committee at the University of Windsor (AUPP 21-13).
Body condition manipulation
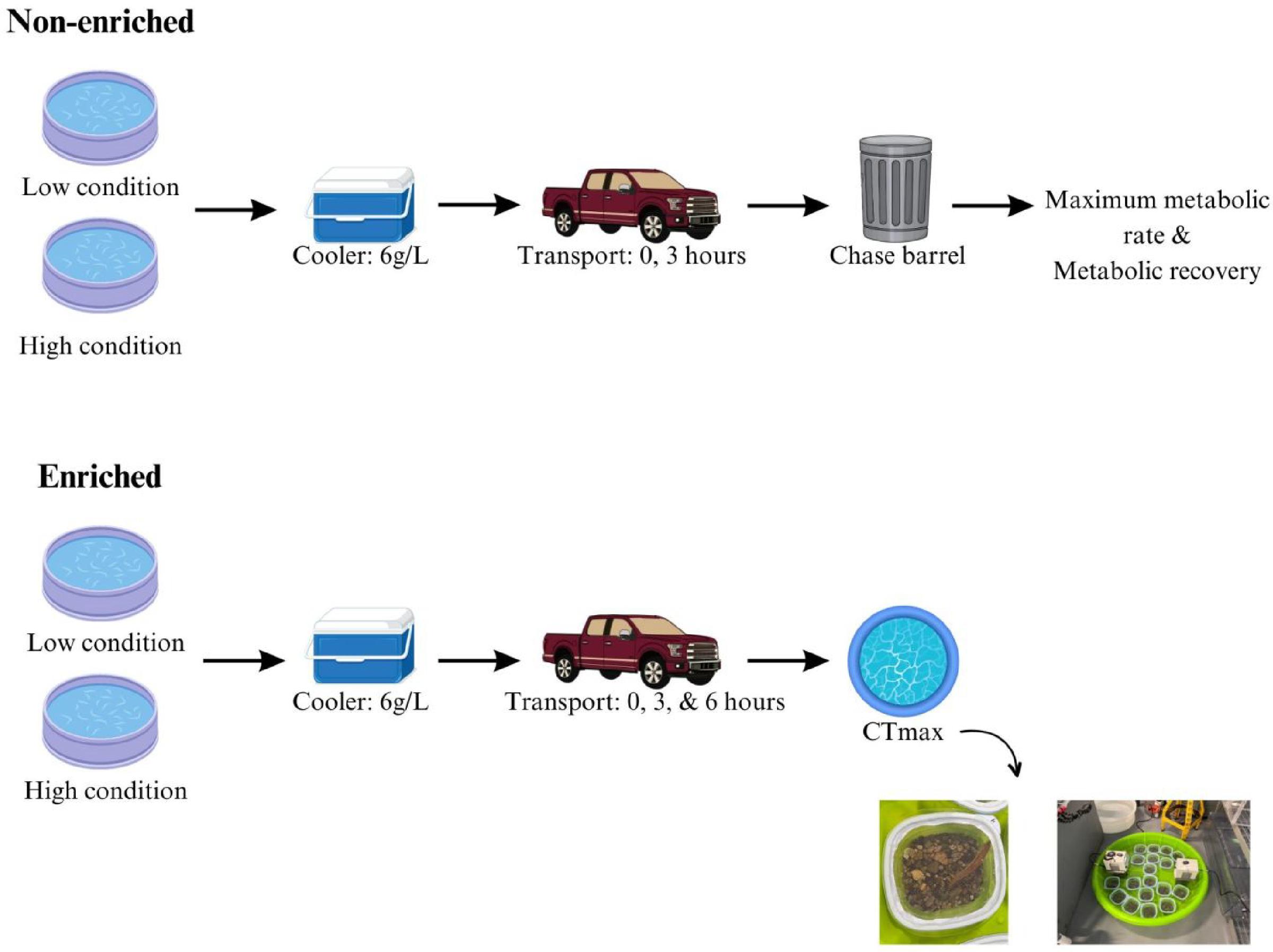
For approximately 12 weeks beginning 28 February 2022 (prior to the start of experimentation), redside dace body condition was manipulated prior to transportation. This 12-week period was selected based on findings from a prior study by Turko et al. (2020), which employed a similar timeframe to manipulate body condition in redside dace using low and high ration diets. During this period, redside dace were divided into two groups: one received a low-ration diet, consisting of frozen bloodworms fed daily, which accounted for approximately 1% of their total body mass (below maintenance) (Turko et al. 2020). The other group received a high-ration diet, also consisting of frozen bloodworms fed daily, but the amount provided was approximately 2% of their total body mass (above maintenance) (Turko et al. 2020). To ensure a balanced representation, we randomly selected one enriched tank (n = 49 fish) and one non-enriched tank (n = 30 fish) for the low-ration diet. Similarly, we assigned the remaining two tanks, one enriched (n = 35) and one non-enriched (n = 38), to the high-ration diet. Feeding was suspended for all tanks 24 h prior to transportation, as is common practice prior to transport of captive fishes to decrease waste output and avoid ammonia toxicity (Harmon 2009).
Fish transportation
Transportation trials took place on two consecutive days: 25 May 2022 and 26 May 2022. Each trial day began in the morning when fish with low and high body conditions were collected at random from the non-enriched and enriched tanks and placed into identical coolers (Coleman, 17 L) (Fig. 1). Coolers were selected as they are the intended transportation method for reintroduction efforts. The density between trials was maintained at approximately 6 g/L. We specifically kept densities low in comparison to commercial applications to better mimic ideal conditions for the transport of imperilled species (Whiterod 2019). All coolers were loaded into the backseat of a truck and each cooler was fitted with a bubbler and temperature probe (axGear electronic digital water thermometer). The coolers were secured with ratchet straps to the backseat of the truck to ensure minimal movement or tip-over during transport. Subsequently, redside dace were transported for varying durations: 0, 3, or 6 h. These time points were selected to mimic the expected distances between hatchery facilities and potential reintroduction sites for future conservation initiatives. Throughout transportation, fish were driven along four-lane highway-series roads with smooth surfaces. Brief stops were made every hour to monitor fish for their condition and to record water temperature. Over the course of transport, temperatures in the coolers remained within ±0.2 °C from the initial source water. Oxygen saturation levels were maintained above 95% for the duration of the transport events. Weather conditions remained stable across the days of transportation trials: no rainfall, and ambient temperature was 17.7 °C (25 May) and 23.8 °C (26 May). No deaths occurred during transport.
Fig. 1.
Maximum metabolic rate
Transported fish destined for respirometry trials (day one: n = 4 low and n = 4 high body condition; day two: n = 4 low and n = 4 high body condition) were collected haphazardly from the non-enriched tanks. To accommodate sample size and timing constraints associated with the respirometry set-up, fish were only transported for 3 h. Control fish for the respirometry trials (day one: n = 6 low and n = 6 high body condition; day two: n = 8 low and n = 7 high body condition) were randomly selected from the non-enriched tanks and transferred immediately to chase barrels at the start of the control trials to begin the experiment.
MMR was elicited using a standard exhaustive chase protocol (Little et al. 2020; Raby et al. 2020). Briefly, fish were transferred to circular tanks (57 cm diameter) filled with 10 cm of water and immediately chased by hand for 5 min. Preliminary experiments showed this period of time corresponds to when fish become less responsive to being chased. Fish were then netted, air exposed for 10 s, and rapidly (<10 s) sealed in acrylic respirometers (∼170 mL). Maximum post-exercise oxygen uptake was then measured for 5 min using a fiber-optic sensor (FireSting FSO2-4, PyroScience GmbH) placed inline with a recirculating pump (Eheim 1046) that also ensured mixing of water within the respirometer. Next, oxygen uptake was measured over a 60 min recovery period using intermittent flow respirometry (Clark et al. 2013; Nelson 2016). A second “flush” pump was connected to a digital timer which allowed the pump to introduce well-oxygenated water from a reservoir when turned on and allowed for measurement of oxygen uptake when turned off. Each phase (flush/measurement) lasted for 5 min (Table S1).
MMR was calculated as highest rate of oxygen uptake measured over a 60 s interval in the 5 min measurement period immediately post-chase (Little et al. 2020). Metabolic rate during recovery was estimated using the rates of oxygen uptake during each of the six subsequent measurement intervals.
Thermal tolerance
Low and high body condition fish destined for thermal tolerance experiments were randomly selected from the enriched tanks (day one: n = 30 low and n = 30 high body condition; day two: n = 19 low and n = 5 high body condition) and transported for either 0 (control), 3, or 6 h. The control group did not undergo any transportation but was placed into a cooler and left to acclimate for approximately 15 min before thermal tolerance testing. Additionally, fish of other species (i.e., Creek Chub (Semotilus atromaculatus) and Northern Redbelly Dace (Chrosomus eos), n = 12) were added to the high body condition fish coolers on day two to keep the density between trials consistent at approximately 6 g/L.
We evaluated thermal tolerance of redside dace across low and high body condition fish and transportation treatments by determining the critical thermal maximum (CTMax). At the start of each trial, fish were placed into a holding container and left for 15 min to acclimate to starting temperatures before heating began. A 15 -min acclimation period was chosen as previous work by Turko et al. (2020) found no difference in CTMax between adults acclimated for 15 min versus 60 min. During each CTMax trial (six trials in total), fish were placed individually into a clear plastic holding container (Rubbermaid, 750 ml) (Fig. 1). Each container was perforated and was fit with a mesh lid to ensure water could flow through. One cup (∼ 128 g) of gravel was also added to the bottom of each container to secure them in place. During each trial, 20 containers were placed into a large pool (Gracious Living 46″ Econo Pool; 9.1″ H × 46″ L × 46″ W) filled with recirculating water (80 L; average starting temperature 17.4 °C; range = 15.65–18.26 °C). Each trial represented one transportation driving time (3 trials per day: 0, 3, and 6 h). Temperature and dissolved oxygen were continually monitored during each trial. CTMax was measured by steadily increasing the water temperature at a controlled rate (average: 0.32 °C/min; range = 0.27–0.35 °C/min). Water temperature was controlled by an immersion circulating heater (SC100 Immersion Circulators: Thermo Fisher Scientific) (Warriner et al. 2020). At CTMax (i.e., when fish could no longer swim upright for approximately 3 s), time and temperature (°C) were recorded. Fish were then removed from the water bath and placed into a recovery cooler. After 15 min in recovery, fish were weighed (g) and measured (fork length, cm) before being placed into a separate holding tank designated for recovery. Recovery was monitored for 48 h following the trials. No deaths were observed after the trials concluded.
Data and statistical analyses
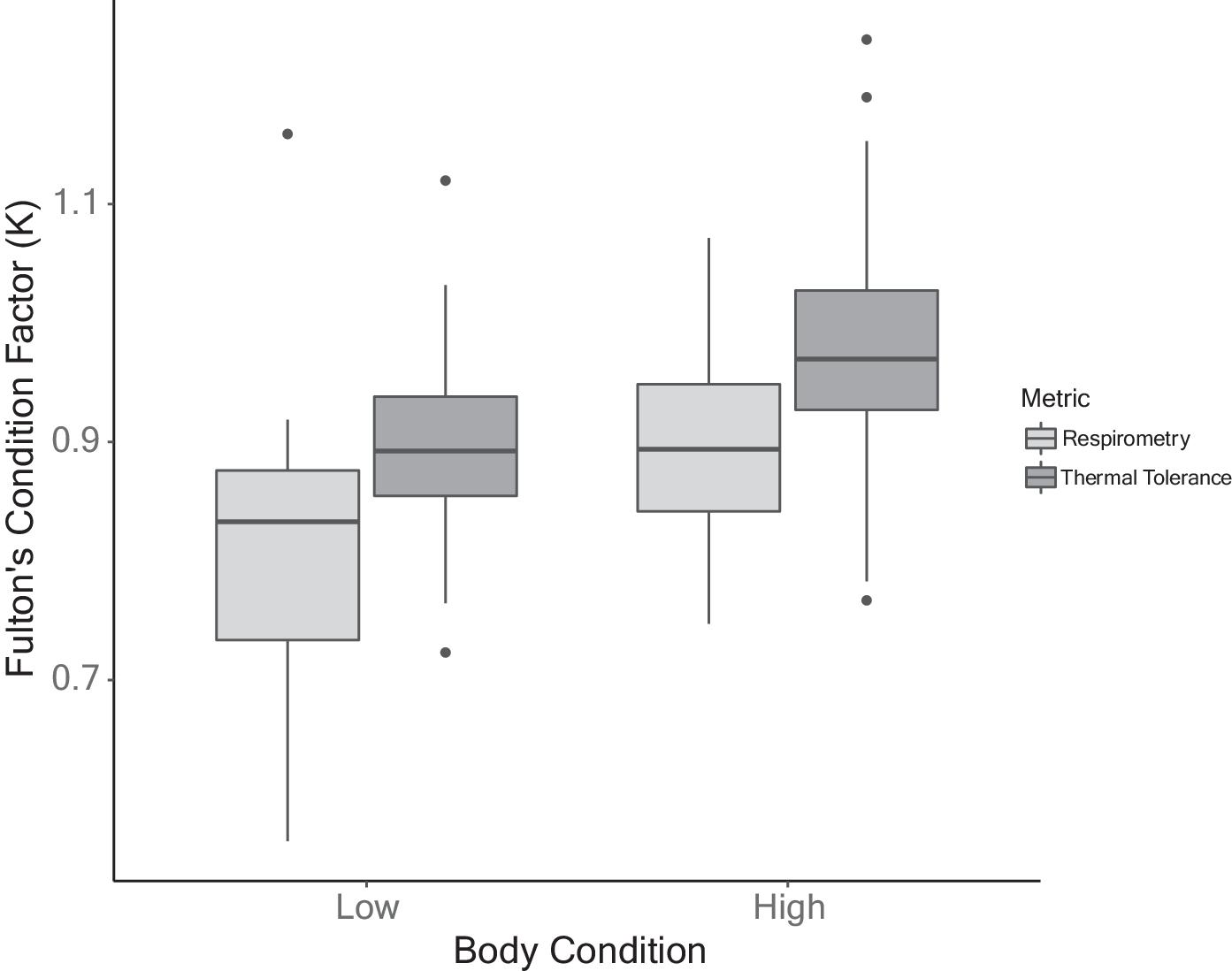
Body condition was assessed from mass (M) and fork length (L) using Fulton’s condition factor (K = M/L3 × 100). We opted for this measurement for its comparability with previous studies (e.g., Turko et al. 2020). To explore the data, t-tests were performed to examine differences in body condition (low vs. high ration) between fish subjected to MMR and thermal tolerance testing. Fish subjected to MMR had a low body condition of 0.80 ± 0.13 and a high body condition of 0.89 ± 0.10 (t-test: t = −2.41, p = 0.02; Fig. 2). Fish used for thermal tolerance testing had slightly higher body condition overall; low ration fish had a body condition of 0.89 ± 0.07 and high ration fish condition was 0.98 ± 0.11 (t-test: t = −4.54, p < 0.0001; Fig. 2).
Fig. 2.
We evaluated the effect of transport and body condition on MMR using a linear mixed effects model (LMM) (R package “nlme”; R Core Development Team 2018). The model examined the association between the dependent variable (MMR) and three independent variables: treatment (control or transported), condition (low or high ration), and mass, while also considering the interaction between treatment and ration (all fixed effects). Additionally, researcher ID was induced as a random effect to account for any variation introduced by the two different individuals who chased the fish during data collection. We conducted a second linear mixed effect model to explore the relationship between metabolic rate following recovery and the independent variables: treatment (control or transported), condition (low or high ration), mass, time, as well as their interactions. To address individual variation among redside dace, fish ID was included as a random effect in the model. Estimated marginal means were calculated following the analysis to make comparisons of metabolism at different time points to better understand recovery time following transportation. To meet the assumption of normality of residuals, MMR was log transformed prior to analysis.
To examine the effects of transportation and body condition on CTmax, an LMM was performed. The model examined the relationship between the dependent variable, CTmax, and two independent variables, transportation (0, 3, or 6 h) and body condition, while accounting for the influence of a random effect, trial number, which was included to account for potential variability across trials. The model of best fit was identified as including both the independent variables and their interaction term—based on model selection. Model assumptions, including linearity, homoscedasticity, and independence of residuals, were assessed through a graphical examination of residuals. Statistical significance was determined using a threshold of p < 0.05. Data are available upon request.
Results
Maximum metabolic rate
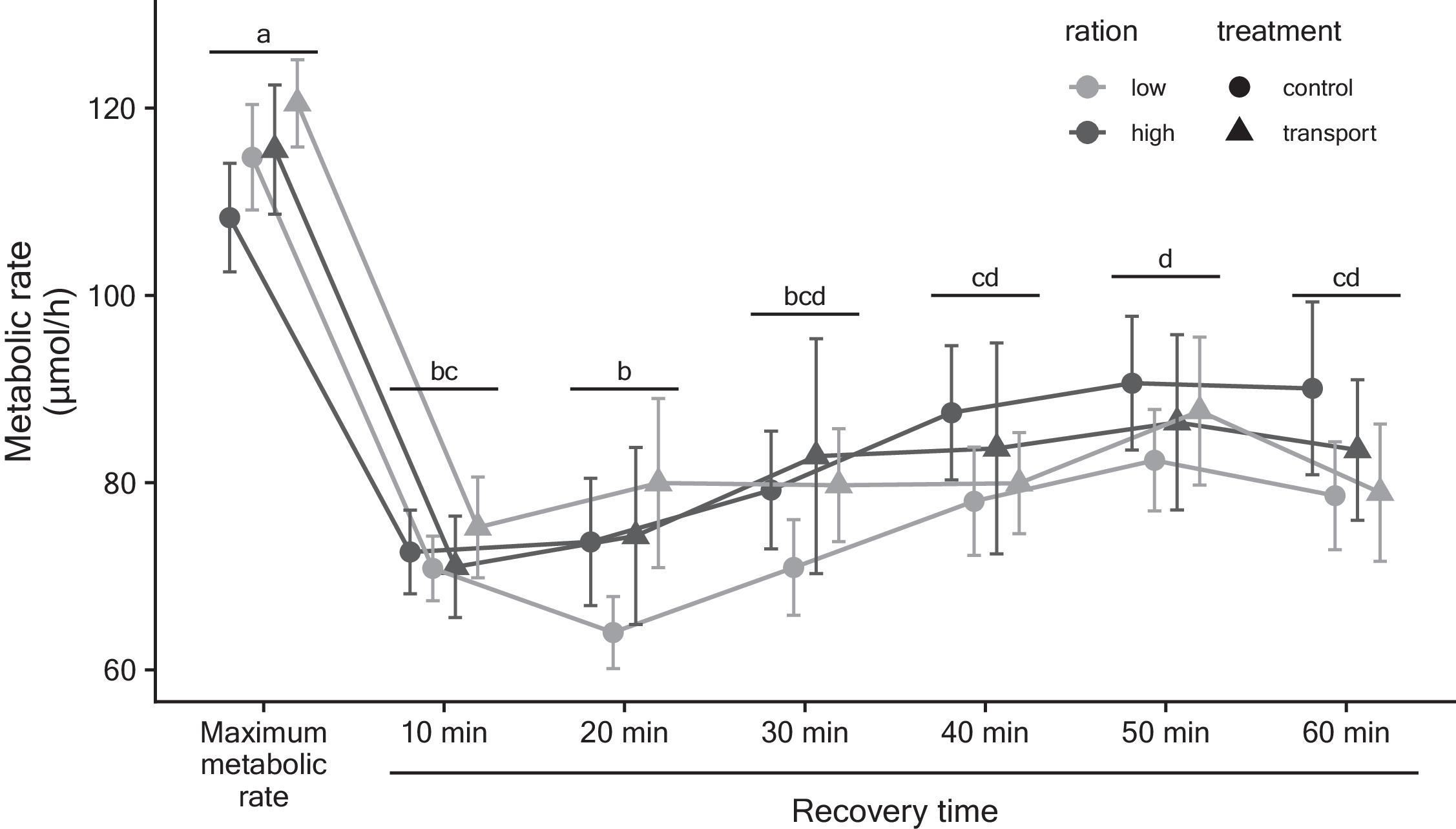
There was no significant effect of transport or condition on MMR (treatment F[1,39] = 0.60, P = 0.44; condition F[1,39] = 0.27, P = 0.61; interaction F[1,39] = 0.03, P = 0.87). Mass was positively correlated with MMR as expected (F[1,39] = 16.13, P = 0.0003). Recovery from exhaustive exercise was also unaffected by transport (F[1,40] = 0.16, P = 0.70), condition (F[1,40] = 0.87, P = 0.36), and the interactions of treatment × condition (F[1,40] = 0.67, P = 0.42), time × treatment (F[1,40] = 0.85, P = 0.53), time × condition (F[1,40] = 1.02, P = 0.41), and time × treatment × condition (F[1,40] = 0.36, P = 0.90). Body mass (F[1,40] = 6.44, P = 0.02) and time (F[1,40] = 40.67, P < 0.0001) were found to significantly effect recovery—metabolic rate was positively related to body mass, and oxygen consumption decreased over time (all p-values < 0.0001; Fig. 3).
Fig. 3.
Thermal tolerance
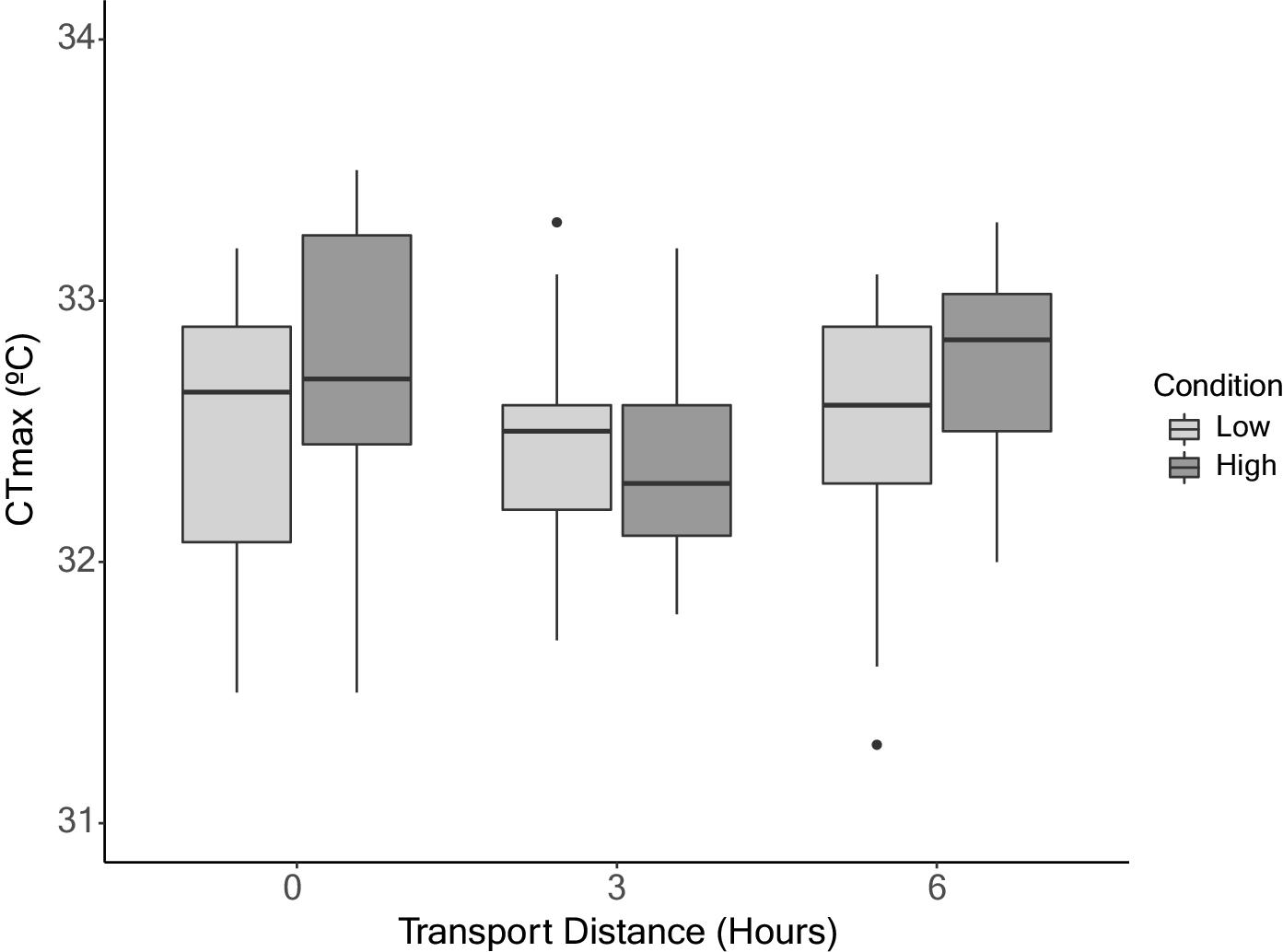
Regardless of transportation distance, no differences in CTmax were observed among redside dace (mean ± standard deviation; 0 h: 32.5 ± 0.6 °C, 3 h: 32.4 ± 0.5 °C, 6 h: 32.7 ± 0.4 °C). Similarly, there were no differences in CTmax between redside dace of low body condition (32.5 ± 0.5 °C) and high body condition (32.6 ± 0.6 °C). The LMM revealed transportation (F[2,3] = 0.40, P = 0.70), body condition (F[1,75] = 0.63, P = 0.43), and their interaction (F[2,75] = 0.06, P = 0.94) did not have a significant effect on CTmax (Fig. 4).
Fig. 4.
Discussion and conclusions
We found that MMR, metabolic recovery, and thermal tolerance did not differ between transported and control fish and that body condition had no effect on the response to transport across any metrics investigated. Our study represents an initial assessment of the influence of transport on redside dace physiology, and a preliminary investigation of whether thermal tolerance is altered by transport in a small-bodied fish species. Given the environmental sensitivity of redside dace and the potential for individuals to be exposed to multiple stressors in their native range (Redside Dace Recovery Team 2010; Turko et al. 2020), this research has implications for the recovery and reintroduction planning in this species. Our findings bode well for reintroduction efforts of redside dace, as the results of our study indicate that transport does not significantly affect the fitness-related endpoints we examined. However, wild/in-stream assessments of fitness are necessary to validate this assumption.
MMR was neither influenced by transport nor body condition (low vs. high ration) in our captive population of redside dace. Variation in MMR within a species has been shown to correlate with intrinsic and extrinsic factors including cardiac function, swimming performance, and the ability to flexibly respond to environmental perturbations (Norin and Clark 2016). In addition, there is some evidence that oxygen consumption may reach values in wild fish that is close to their MMR while undertaking normal daily activities (Murchie et al. 2011; Norin and Clark 2016), further reinforcing the value of this physiological metric for understanding whether fish have the capacity to cope with different types of changes in their environment. In the case of our experiment, the capture, handling, and chase protocol may have been the largest contributor to MMR (Davis and Shreck 1997), with the transport event not leading to any change or impairment to maximum aerobic capacity. Similarly, juvenile spotted grunters (Pomodasys comersonnii) displayed the greatest metabolic response to capture and chase, with the response to additional stressors of acute temperature increase and high packing density not leading to lower MMR in comparison (Radull et al. 2002). The transport time we employed was relatively short at 3 h and may have represented a transient demand on experimental fish. Longer transport times are routinely used in other fish species and could lead to impacts on MMR.
Interestingly, although the food supplementation increased body condition, it did not result in differences in MMR between supplemented and control fish. This finding is contrary to some previously reported results where improved body condition lowered MMR. For example, active metabolic rate of Atlantic Cod (Gadus morhua L.) decreased after a 9-week feeding regime that resulted in higher condition factor compared to condition at the time of capture from the wild (Lapointe et al. 2006). However, other studies have found results similar to ours; for instance, MMR of Brown Trout (Salmo trutta) did not change in relation to increasing food availability (low, intermediate, ad libitum) (Auer et al. 2016). It is possible that given, that the fish used in our experiments had been in a long-term hatchery setting and our treatments did not create a food shortage, the condition of all fish was high enough that MMR was unaffected. Indeed, it has been proposed that the predominant contributor to MMR may be skeletal muscle, which would have been unaffected by our ration treatments, and would be more likely to be affected by very limited food availability or prolonged starvation conditions (Weibel et al. 2004; Auer et al. 2016).
Metabolic recovery did not differ between transported and non-transported fish and was also not influenced by body condition. Under other types of stressors for fishes, recovery rate has been shown to be similar across stressor-exposed and control groups. For example, Powell et al. (2005) found that post-exercise recovery of metabolic rate was the same in healthy Rainbow Trout compared to those infected with the microsporidium gill parasite Loma salmonae. Further, through an indirect assessment of metabolic recovery rate using heart rate in Sockeye Salmon (Oncorhynchus nerka), the recovery time of fish that underwent exhaustive exercise did not differ from those that underwent exhaustive exercise and were also exposed to predator alarm cue (Lawrence et al. 2023). Given that recovery metabolism can place constraints on overall performance (Milligan 1996) including the locomotor capacity to evade predation (Cooke et al. 2014), our finding that the transport event did not result in an altered recovery trajectory in comparison to control fish is encouraging for post-release recovery of redside dace.
CTmax was also not influenced by transportation and body condition. While the effect of transport on CTmax in small-bodied fish remains unexplored, our findings suggest that redside dace are robust to the acute stressors associated with reintroduction. Greater body condition and/or dietary enrichment have also been associated with increased thermal tolerance in other fishes (e.g., Robinson et al. 2008; Tejpal et al. 2014; Lee et al. 2016; Turko et al. 2020). This relationship is thought to be driven by increased energy reserves, facilitating mitochondrial function, making it possible for fish to meet the energy demands required to cope with physiological stressors, such as elevated temperatures (Ern et al. 2023). However, contrasting findings have emerged regarding the thermal tolerance of larger individuals, which may struggle with increasing temperatures due to their surface-to-mass ratio (Ern et al. 2023). Interestingly, our study did not uncover evidence supporting either association. Identifying a single reason for the lack of influence on CTmax by both transport and body condition remains difficult; however, our attempt to establish optimal transport conditions provides valuable insights. In this study, we aimed to mimic the priorities that would be in place during the transport and reintroduction process of an endangered species. As a result, fish were transported under low density (6 g/L) and in the climate-controlled section of a vehicle, allowing us to maintain very stable oxygen and temperature conditions. The timeframe of 3–6 h was chosen to align with likely travel distances for a reintroduction program for this species and would greatly decrease the potential for changes in water quality parameters (e.g., ammonia) (Sampaio and Freire 2016). Altogether, these transport conditions may not have been harsh enough to elicit a marked difference in CTmax or even MMR or recovery (see above) in comparison to control (non-transported) fish. Further, the additional energetic stores of food-supplemented fish would not impart a benefit in these conditions because all fish, regardless of treatment, were in high enough condition to maintain optimal physiological functions. It is possible that during transport, food supplementation may still be a useful technique for minimizing transport stress, and we suggest that future investigations continue to employ transport times that are relevant for species-specific transport and release protocols.
Overall, our study aimed to assess the effect of transport on fitness-related metrics relevant to reintroduction, specifically MMR and CTmax. We also examined supplemental feeding to mitigate these effects. Our results show that neither transport nor body condition significantly influenced MMR, recovery, or CTmax in our captive redside dace population. These findings lend preliminary support to the viability of successfully transporting redside dace from captivity to reintroduction sites. Future research could extend this work by including fish transported from natural habitats, thereby offering insights that could guide timing decisions. While our study did not find an impact of transport on metabolism and thermal tolerance, behavioral traits, such as food intake, risk-taking behavior, orientation, and activity level, could also impact survival in a release site and should be investigated in the future. It is however encouraging that a transport event designed to simulate best-practices for this species did not result in negative effects for MMR, metabolic recovery, or thermal tolerance, given that these aspects of physiological functioning would also be important for persistence in a novel environment. In conclusion, our study provides key insight into the interplay between transportation stress, metabolism, and thermal tolerance in redside dace. Taken together, our findings underscore the feasibility of transporting redside dace to a release site and this information may possibly be extended to other small-bodied fish, for which information is lacking.
Acknowledgements
We thank Dane Roberts, who helped transport redside dace. The research was funded by a Natural Sciences and Engineering Research Council CREATE grant (FishCAST), the Municipality of Peel Region and Fisheries and Oceans Canada.
References
Auer S.K., Salin K., Rudolf A.M., Anderson G.J., Metcalfe N.B. 2016. Differential effects of food availability on minimum and maximum rates of metabolism. Biology Letters, 12(10): 20160586.
Barton B.A., Morgan J.D., Vijayan M.M. 2002. Physiological and condition-related indicators of environmental stress in fish. In Biological indicators of aquatic ecosystem stress. Edited by S.M. Adams. American Fisheries Society. pp. 111–148.
Clark T.D., Sandblom E., Jutfelt F. 2013. Aerobic scope measurements of fishes in an era of climate change: respirometry, relevance and recommendations. The Journal of Experimental Biology, 216(15): 2771–2782.
Cochran-Biederman J.L., Wyman K.E., French W.E., Loppnow G.L. 2015. Identifying correlates of success and failure of native freshwater fish reintroductions. Conservation Biology, 29(1): 175–186.
Cooke S.J., Messmer V., Tobin A.J., Pratchett M.S., Clark T.D. 2014. Refuge-seeking impairments mirror metabolic recovery following fisheries-related stressors in the Spanish flag snapper (Lutjanus carponotatus) on the Great Barrier Reef. Physiological and Biochemical Zoology, 87(1): 136–147.
COSEWIC. 2017. COSEWIC assessment and status report on redside dace clinostomus elongatus in Canada. Ottawa: Committee on the status of endangered wildlife in Canada. p. 63.
Davis L.E., Schreck C.B. 1997. The energetic response to handling stress in juvenile coho salmon. Transactions of the American Fisheries Society, 126(2): 248–258.
Dickens M.J., Delehanty D.J., Romero L.M. 2010. Stress: an inevitable component of animal translocation. Biology Conservation, 143(6): 1329–1341.
Drake D.A.R., Lamothe K.A., Thiessen K.E., Morris T.J., Koops M.A., Pratt T.C., et al. 2021. Fifteen years of Canada's Species at Risk Act: evaluating research progress for aquatic species in the Great Lakes—St. Lawrence River basin. Canadian Journl of Fisheries and Aquatic Sciences, 78(9): 1205–1218.
Ern R., Andreassen A.H., Jutfelt F. 2023. Physiological mechanisms of acute upper thermal tolerance in fish. Physiology, 38(3): 141–158.
Fry F.E.J. 1971. The effect of environmental factors on the physiology of fish. In Fish physiology. Edited by W.S. Hoar, D.J. Randall. Academic Press, Inc. pp. 1–98.
Harmon T.S. 2009. Methods for reducing stressors and maintaining water quality associated with live fish transport in tanks: a review of the basics. Reviews in Aquaculture, 1(1): 58–66.
Killen S.S., Norin T., Halsey L.G. 2017. Do method and species lifestyle affect measures of maximum metabolic rate in fishes? Journal of Fish Biology, 90(3): 1037–1046.
Lapointe D., Guderley H., Dutil J.D. 2006. Changes in the condition factor have an impact on metabolic rate and swimming performance relationships in Atlantic cod (Gadus morhua L.). Physiological and Biochemical Zoology, 79(1): 109–119.
Lamothe K.A., Drake D.A.R., Pitcher T.E., Broome J.E., Dextrase A.J., Gillespie A., et al. 2019. Reintroduction of fishes in Canada: a review of research progress for SARA-listed species. Environmental Reviews, 27(4): 575–599.
Lawrence M.J., Prystay T.S., Dick M., Eliason E.J., Elvidge C.K., Hinch S.G., et al. 2023. Metabolic constraints and individual variation shape the trade-off between physiological recovery and antipredator responses in adult sockeye salmon. Journal of Fish Biology, 103(2): 280–291.
LeBlanc S., Middleton S., Gilmour K.M., Currie S. 2011. Chronic social stress impairs thermal tolerance in the rainbow trout (Oncorhynchus mykiss). The Journal of Experimental Biology, 214(10): 1721–1731.
Lee S., Hung S.S., Fangue N.A., Haller L., Verhille C.E., Zhao J., Todgham A.E. 2016. Effects of feed restriction on the upper temperature tolerance and heat shock response in juvenile green and white sturgeon. Comparitive Biochemistry and Physiology. Part A, Molecular and Integrative Physiology, 198: 87–95.
Little A.G., Dressler T., Kraskura K., Hardison E., Hendriks B., Prystay T., et al. 2020. Maxed out: optimizing accuracy, precision, and power for field measures of maximum metabolic rate in fishes. Physiological and Biochemical Zoology, 93(3): 243–254.
Lutterschmidt W.I., Hutchison V.H. 1997. The critical thermal maximum: history and critique. Canadian Journal of Zoology, 75: 1561–1574.
McEwen B.S., Wingfield J.C. 2003. The concept of allostasis in biology and biomedicine. Hormones and Behavior, 43(1): 2–15.
Milligan C.L. 1996. Metabolic recovery from exhaustive exercise in rainbow trout. Comparitive Biochemistry and Physiology. Part A, Physiology, 113(1): 51–60.
Monirian J., Sutphin Z., Myrick C. 2010. Effects of holding temperatures and handling stress on the upper thermal tolerance of threadfin shad Dorosoma petenense. Journal of Fish Biology, 76(6): 1329–1342.
Murchie K.J., Cooke S.J., Danylchuk A.J., Suski C.D. 2011. Estimates of field activity and metabolic rates of bonefish (Albula vulpes) in coastal marine habitats using acoustic tri-axial accelerometer transmitters and intermittent-flow respirometry. Journal of Experimental Marine Biology and Ecology, 396(2): 147–155.
Näslund J. 2021. Reared to become wild-like: addressing behavioral and cognitive deficits in cultured aquatic animals destined for stocking into natural environments—a critical review. Bulletin of Marine Science, 97(4): 489–538.
Nelson J.A. 2016. Oxygen consumption rate v. rate of energy utilization of fishes: a comparison and brief history of the two measurements. Journal of Fish Biology, 88(1): 10–25.
Norin T., Clark T.D. 2016. Measurement and relevance of maximum metabolic rate in fishes. Journal of Fish Biology, 88(1): 122–151.
Parker B.J., McKee P., Campbell R.R. 1988. Status of the redside dace, Clinostomus elongatus, in Canada. The Canadian Field-Naturalist, 102: 163–169.
Pörtner H.O., Peck M.A. 2010. Climate change effects on fishes and fisheries: towards a cause-and-effect understanding. Journal of Fish Biology, 77(8): 1745–1779.
Powell M.D., Speare D.J., Daley J., Lovy J. 2005. Differences in metabolic response to Loma salmonae infection in juvenile rainbow trout oncorhynchus mykiss and brook trout Salvelinus fontinalis. Disease of Aquatic Organisms, 67: 233–237.
R Development Core Team. 2018. R: a language and environment for statistical computing. Foundation for Statistical Computing. Available from www.R-project.org [accessed 6 July 2022].
Raby G.D., Doherty C.L.J., Mokdad A., Pitcher T.E., Fisk A.T. 2020. Post-exercise respirometry underestimates maximum metabolic rate in juvenile salmon. Conservation Physiology, 8: coaa063.
Radull J., Kaiser H., Hecht T. 2002. Stress-related changes in the metabolic rate of juvenile spotted grunter, Pomadasys commersonnii (Haemulidae, Pisces). Marine Freshwater Research, 53(2): 465–469.
Redside Dace Recovery Team. 2010. Recovery strategy for redside dace (Clinostomus elongatus) in Ontario. Ontario recovery strategy series. p. 29.
Robinson M.L., Gomez-Raya L., Rauw W.M., Peacock M.M. 2008. Fulton's body condition factor K correlates with survival time in a thermal challenge experiment in juvenile Lahontan cutthroat trout (Oncorhynchus clarki henshawi). Journal of Thermal Biology, 33(6): 363–368.
Sampaio F.D.F., Freire C.A. 2016. An overview of stress physiology of fish transport: changes in water quality as a function of transport duration. Fish and Fisheries, 17(4): 1055–1072.
Schulte P.M., Healy T.M., Fangue N.A. 2011. Thermal performance curves, phenotypic plasticity, and the time scales of temperature exposure. Integrative and Comparitive Biology, 51(5): 691–702.
Sébastien A., Manuel G., Bastien S. 2021. Temperature increases and its effects on fish stress physiology in the context of global warming. Journal of Fish Biology, 98(6): 1496–1508.
Seddon P.J., Armstrong D.P., Maloney R.F. 2007. Developing the science of reintroduction biology. Conservation Biology, 21(2): 303–312.
Seddon P.J., Strauss W.M., Innes J. 2012. Animal transloctions: what are they and why do we do them? In Reintroduction biology: integrating science and management. Edited by J.G. Ewen, D.P. Armstrong, K.A. Parker, P.J. Seddon. Wiley-Blackwell. pp.1–32.
Serrao N.R., Reid S.M., Wilson C.C. 2018. Conservation genetics of redside dace (Clinostomus elongatus): phylogeography and contemporary spatial structure. Conservation Genetics, 19: 409–424.
Teixeira C.P., Schetini de Azevedo C., Mendl M., Cipreste C.F., Young R.J. 2007. Revisiting translocation and reintroduction programmes: the importance of considering stress. Animal Behavior, 73(1): 1–13.
Tejpal C.S., Sumitha E.B., Pal A.K., Shivananda M.H., Sahu N.P., Siddiah G.M. 2014. Effect of dietary supplementation of L-tryptophan on thermal tolerance and oxygen consumption rate in Cirrhinus mrigala fingerlings under varied stocking density. Journal of Thermal Biology, 41: 59–64.
Turko A.J., Nolan C.B., Balshine S., Scott G.R., Pitcher T.E. 2020. Thermal tolerance depends on season, age and body condition in imperilled redside dace Clinostomus elongatus. Conservation Physiology, 8(1): 1–15.
Vanderzwalmen M., Eaton L., Mullen C., Henriquez F., Carey P., Snellgrove D., Sloman K.A. 2019. The use of feed and water additives for live fish transport. Reviews in Aquaculture, 11(1): 263–278.
Warriner T.R., Semeniuk C.A.D., Pitcher T.E., Heath D.D., Love O.P. 2020. Mimicking transgenerational signals of future stress: thermal tolerance of juvenile Chinook Salmon is more sensitive to elevated rearing temperature than exogenously increased egg cortisol. Frontiers in Ecology and Evolution, 8: 548939.
Weibel E.R., Bacigalupe L.D., Schmitt B., Hoppeler H. 2004. Allometric scaling of maximal metabolic rate in mammals: muscle aerobic capacity as determinant factor. Respiratory Physiology and Neurobiology, 140(2): 115–132.
Wentworth C.K. 1922. A scale of grade and class terms for clastic sediments. The Journal of Geology, 30(5): 377–392.
Whiterod N. 2019. A translocation strategy to ensure the long-term future of threatened small-bodied freshwater fishes in the South Australian section of the Murray-Darling Basin: a report to Natural Resources, SA Murray-Darling Basin and the Riverine Recovery Project. Aquasave–Nature Glenelg Trust. 97p.
Supplementary material
Supplementary Material 1 (DOCX / 22.9 KB).
- Download
- 22.99 KB
Information & Authors
Information
Published In

FACETS
Volume 9 • Number 1 • January 2024
Pages: 1 - 9
Editor: Marco Rodriguez
History
Received: 24 November 2023
Accepted: 23 April 2024
Version of record online: 14 August 2024
Notes
This paper is part of a collection entitled Progress and Priorities for the Recovery of Aquatic Species at Risk in Canada.
Copyright
© 2024 The Author(s). This work is licensed under a Creative Commons Attribution 4.0 International License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Data Availability Statement
Data generated or analyzed during this study are available from the corresponding author upon reasonable request.
Key Words
Sections
Subjects
Authors
Author Contributions
Conceptualization: AMW, CLM, AJT, AIM, TEP
Formal analysis: AMW, AJT
Funding acquisition: TEP
Investigation: AMW, CLM, AJT, AIM
Methodology: AMW, CLM, AJT, TEP
Project administration: AMW, CLM, AJT, AIM
Resources: AJT, TEP
Supervision: AJT, TEP
Writing – original draft: AMW, CLM
Writing – review & editing: AJT, AIM, TEP
Competing Interests
The authors declare there are no competing interests.
Metrics & Citations
Metrics
Other Metrics
Citations
Cite As
Ashley M. Watt, Christine L. Madliger, Andy J. Turko, Ali I. Mokdad, and Trevor E. Pitcher. 2024. Transport does not influence maximum metabolic rate or thermal tolerance of endangered redside dace (Clinostomus elongatus). FACETS.
9: 1-9.
https://doi.org/10.1139/facets-2023-0206
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
There are no citations for this item