The effect of riffle restoration on the recovery of endangered Nooksack Dace (Rhinichthys cataractae sp. cataractae)
Abstract
The Nooksack Dace (Rhinichthys cataractae sp. cataractae) is a federally endangered riffle specialist endemic to the lower Fraser Valley of British Columbia, Canada, with historic population declines associated with riffle loss from stream dredging, channelization, and excessive sediment inputs. To assess the effectiveness of riffle restoration as a recovery strategy, gravel and cobble riffles were constructed in two replicate tributaries of the Nooksack River as a before-after-control-impact experiment, measuring dace abundance, substrate composition, and invertebrate biomass before and one year after restoration. Nooksack Dace density increased significantly in cobble (but not gravel) treatments relative to control riffles. Dace abundance was strongly associated with increased availability of interstitial refuges rather than substrate effects on invertebrate prey abundance, suggesting that interstitial space limits adult dace abundance. Young-of-the-year dace were not observed in one of the two restored streams despite riffle restoration, indicating increased dace density due to aggregation in higher-quality restored riffles. This recruitment limitation indicates persistence of a population bottleneck at an early life history stage that is not addressed by successful restoration of adult riffle habitat.
Introduction
Freshwater ecosystems cover less than 1% of the earth’s surface and are inhabited by approximately 40% of global fish biodiversity, but represent some of the most threatened habitats in the world (Dudgeon et al. 2006; Göthe et al. 2015). Habitat loss and degradation, over exploitation, pollution, and flow modification represent the greatest threats to freshwater ecosystems, all of which are directly associated with anthropogenic activities and development (Dudgeon et al. 2006; Arthington et al. 2016; Reid et al. 2019). The deterioration and destruction of freshwater habitats highlight why freshwater fishes are some of the most imperiled organisms in the world (Duncan and Lockwood 2001; Jelks et al. 2008).
Although up to 30% of freshwater fishes are threatened with extinction (Collen et al. 2014; Miranda et al. 2022), not all trophic guilds are equally vulnerable to habitat degradation. Elevated sediment input is likely the single most common aspect of degraded stream habitat (Kemp et al. 2011); consequently, the guild of benthic insectivores are particularly vulnerable to watershed development, because their habitat and prey resources are directly impacted by sediment deposition (Rabeni and Smale 1995; Walters et al. 2003). This vulnerability is compounded for riffle-dependent benthic insectivores with high water velocity requirements (Aadland 1993), since flow reduction is another frequent consequence of human development of stream ecosystems (de Graaf et al. 2019). Additional risk exists if the distribution of a benthic insectivore coincides with human development pressure, and if they have small body size, since small-bodied species are also more susceptible to extinction (Reynolds et al. 2005; Vélez-Espino and Koops 2012; van der Lee and Koops 2015).
The Nooksack Dace (Rhinichthys cataractae sp. cataractae), a small, benthic cyprinid and riffle-specialist insectivore, is a genetically and morphologically distinct form of Longnose Dace (Rhinichthys cataractae) of uncertain taxonomic status (COSEWIC 2007). It diverged from other forms of R. cataractae during geographic isolation in a glacial refuge in Washington State during the Pleistocene (Taylor et al. 2015). Nooksack Dace is endemic to only four watersheds in Canada in the lower Fraser Valley (LFV) of British Columbia (Pearson et al. 2007), where it is listed as Endangered under the federal Species at Risk Act (SARA; Fisheries and Oceans Canada 2020) and by the Committee on the Status of Endangered Wildlife in Canada (COSEWIC 2018). Rapidly declining populations are attributed to a range of impacts including habitat loss and degradation, effects which are compounded by low flows and deteriorating water quality, all resulting from anthropogenic development within their watersheds (Pearson 1998, 1999; Pearson et al. 2007; Boyd et al. 2022).
Dace are typically found in riffle habitat (Pearson et al. 2007), preferring shallow moving water (>0.25 m·s−1) (McPhail 1996) over coarse unembedded substrate. Individuals will forage in pools during drought when riffles dry (Avery-Gomm et al. 2014) and as young-of-the-year when they inhabit shallow, marginal pools with fine substrate adjacent to riffle spawning habitat (McPhail 1996; Pearson 1998, 1999). Dace rely on the interstitial spaces amongst the coarse substrate particles of the bed for refuge from flow and predation, for spawning, and for feeding (Pearson 1999). However, their dependence on riffles with unembedded coarse substrate leaves the species particularly vulnerable to habitat degradation resulting in the deposition of fine sediment.
Similar to many valley bottom floodplain landscapes throughout the world, the LFV has been transformed by human activities over the last 200 years. Here, a mix of temperate rainforest and wetlands became a landscape dominated by agriculture, resource extraction, and increasing urbanization (Boyle et al. 1997; Pearson 1998). In the process, 15% of the streams within the LFV no longer exist, were paved over, or piped (Fisheries and Oceans Canada 1998) with many more channelized and dredged (Pearson et al. 2007). While channelization is beneficial for drainage and flood mitigation, it results in homogenous habitat lacking the physical complexity associated with a natural meandering channel (Rambaud et al. 2009; Lennox III and Rasmussen 2016). This habitat complexity is essential to accommodate the diversity of habitats required by different life history stages for spawning, juvenile rearing, feeding, and hydraulic refuge during floods (Pearson et al. 2007).
In addition to direct habitat simplification through channelization, alteration of upslope processes by agriculture and urbanization typically increase fine sediment inputs to streams (Kemp et al. 2011). Although the transport and storage of organic and inorganic sediments is an integral process driving substrate heterogeneity and nutrient transport, excessive deposition of fine sediment alters channel structure and substrate properties (i.e., particle size, interstitial volume, and hyporheic flow) in ways that are often detrimental to fish and invertebrate production (Wood and Armitage 1997; Florsheim et al. 2008; Scholl et al. 2022). Urban storm run-off, agricultural effluent, and accelerated bank erosion all increase the amount and frequency of sediment inputs (Suttle et al. 2004).
Infilling of interstitial space by fine sediment inputs directly removes foraging and refuge habitat for benthic fish like Nooksack Dace, but can also limit the interstitial flow of oxygen rich water to developing eggs and larvae (Pearson et al. 2007; Kemp et al. 2011; Kawanishi et al. 2015). Excessive fines can also decrease benthic invertebrate diversity and production by clogging substrate interstices (Bo et al. 2007; Burdon et al. 2013), an effect that is particularly pronounced in small lowland streams that lack the hydraulic power to export deposited fines (Naden et al. 2016). The unembedded substrate of riffle habitats represent hotspots of benthic invertebrate production in streams (Poff and Huryn 1998; Scholl et al. 2022). Infilling of interstitial space can greatly diminish prey abundance for fish (Suttle et al. 2004), in addition to reducing space available for refuging (Finstad et al. 2007), providing both direct and indirect pathways for interstitial accumulation of fine sediment to impact fish.
Riffle habitat restoration is a practical approach to recover endangered riffle specialists, but its effectiveness and underlying pathways of effect are rarely rigorously tested. To assess the effectiveness of riffle habitat restoration at increasing abundance of a typical riffle benthivore, we added coarse unembedded gravel and cobble substrate to existing riffles in two streams with contrasting riffle habitat quality: one with unembedded substrate and high dace densities, and a second with scarce coarse substrate and few dace. The main objectives were to evaluate the overall effectiveness of coarse substrate addition at increasing dace abundance, and to identify the specific processes and pathways that lead to increased density. Our expectations were that any change in dace abundance following substrate addition would be most pronounced in the stream with more degraded riffles, and minimal in the stream with abundant coarse substrate. We executed a before-after-control-impact (BACI; Green 1979) experiment to determine the outcome of gravel and cobble additions to habitat, invertebrates, and fish in two representative Nooksack River tributaries, as described below. We specifically hypothesized that dace abundance would be correlated with the volume of available interstitial space, which would be positively associated with mean particle size and would increase with substrate addition. We also predicted that benthic invertebrate biomass would increase with the availability of interstitial spaces, and would correlate with observed dace density. Finally, we expected a positive response of Nooksack Dace to coarse substrate addition both in terms of greater adult density and increased spawning success (measured in terms of more juvenile recruits following substrate addition).
Materials and methods
Study site and species
Experiments were performed in Bertrand and Pepin Creeks, two transboundary tributaries of the Nooksack River that flow south from British Columbia into Washington State, United States (Fig. S1). Substrate was added to riffles distributed over approximately 1 km of stream channel in both systems (Tables S1 and S2). Bertrand Creek and Pepin Creek both have gradients of approximately 1%–2% and summer low flow wetted widths of approximately 3 m. The main habitat contrast between streams is that Bertrand Creek has a higher proportion of unembedded cobble substrate and abundant dace (1.4 dace·m−2 in pre-treatment sampling), while cobble substrate is scarce and embedded in Pepin Creek, and dace density is low (0.2 dace·m−2 in pre-treatment sampling). Bertrand Creek experiences higher peak flows than Pepin Creek (42 m3·s−1 vs. 1.5 m3·s−1, respectively; Pearson 2004; U.S. Geological Survey 2023) which creates a more mobile channel that more effectively exports fine sediment than the more stable Pepin Creek. However, Bertrand Creek also experiences lower summer minimum flows (0.02 m3·s−1 vs. 0.07 m3·s−1, respectively) because Pepin Creek has a greater groundwater influence (Berg and Allen 2007).
Study design
The substrate composition of control riffles was typically coarse gravel and cobble substrate embedded with accumulated fine substrate and sand (Fig. 1a), but differed between streams. In Pepin Creek, control riffles tended to contain less cobble substrate cover and smaller particles that were more embedded. In comparison, control riffles in Bertrand Creek tended to contain more cobble substrate which tended to be less embedded, likely due to higher peak flows.
Fig. 1.

Restoration experiments included four treatments: control riffles that were unmanipulated, riffles with cobble substrate addition, riffles with gravel substrate addition, and riffles with both cobble and gravel addition as discrete adjacent patches; replicate substrate patches were 3 m long, with the exception of the combined cobble and gravel treatments that were 6 m long and included two sequential patches of gravel and cobble placed in longer riffles. This treatment was intended to assess whether close proximity to unembedded gravel substrate with potentially high benthic invertebrate abundance would increase Nooksack Dace density in adjacent cobble substrate. More treatments were placed in Pepin Creek where lower dace abundance and poorer substrate quality was expected to elicit a larger and more informative response to riffle restoration than in Bertrand Creek, where dace and cobble were already abundant. However, the number and location of replicate riffles was ultimately limited by resources and site access. Three replicates of each of the gravel, cobble, combined cobble-gravel, and control treatments were installed in Pepin Creek (n = 12). However, while all four treatments were installed in Pepin Creek, site and resource constraints limited treatments in Bertrand Creek to only four cobble and four control replicates (n = 8). The assignment of treatments to riffles was randomized to the extent possible however, the location of some treatment replicates was partially constrained by riffle length and site access (i.e., some controls were assigned to riffles where substrate could not be added due to poor machine access). Because substrate-specific dace density in the treatment with adjacent cobble-gravel patches did not differ from density in the isolated gravel and cobble treatments, the combined treatment was merged with the gravel and cobble addition treatment replicates in post-restoration monitoring and assessment to simplify data analysis. Thus, post-restoration analysis included three control replicates and six replicates of both the gravel and cobble treatments in Pepin Creek (n = 15), and four replicates of control and cobble treatments in Bertrand Creek (n = 8).
The experiment was designed as a BACI study (Green 1979), with sampling in all riffle replicates taking place in June to August 2021 prior to riffle restoration, and in June to August 2022, one year post-restoration. Response variables collected before and after substrate addition included substrate characteristics (i.e., particle size, percentage cover in different substrate classes, and interstitial volume) as well as benthic invertebrate and fish density, as described below.
Substrate was added to riffles in August 2021 during the seasonal low flow instream work window, after fish were removed by multi-pass electrofishing prior to habitat manipulation. Both gravel (2–5 cm b-axis length) and cobble (8–30 cm b-axis length) were added to riffles on top of existing substrate in uniform patches measuring approximately 4 m in length and spanning the approximately 3 m wetted width of the creek, for a total surface area of approximately 12 m2 per riffle. Because both streams were small (approximately three meter wetted width at low flow) this also covered the entire length of many riffles (Fig. 1b).
Substrate was added using a track excavator where possible, or by hand when access to the creek was limited by a full riparian buffer. Cobble substrate patches were established by adding a layer of smaller cobbles (i.e., 8–15 cm b-axis length) over the surface of existing riffle substrate, and then placing larger cobble (i.e., 16–30 cm b-axis length) on top to a total depth of 12–15 cm. Gravel patches were established through addition of a uniform layer of gravel substrate to a depth of approximately 6–10 cm. Record fall floods in 2021 post-restoration settled (re-arranged) substrate in a more natural configuration with minimal export of substrate beyond each treatment patch.
Measuring riffle habitat features
Surface substrate composition of riffles was measured before (July to August 2021) and after (July to August 2022) habitat restoration using modified Wolman Pebble Counts (Wolman 1954). Particle diameter was measured at 25 locations on each of four 40 by 40 cm grids systematically placed along transects across each riffle. Each grid consisted of a 40 by 40 cm polyvinyl chloride pipe frame divided with string into 36 equal-area squares, with substrate particles selected at grid intersections.
Substrate particles were measured along the b-axis (Clapcott et al. 2011) and categorized into five size classes based on a modified Wentworth scale; fines (<0.25 mm), sand (0.25–2 mm), gravel (2–64 mm), cobble (64–256 mm), and boulders (>256 mm; Wentworth 1922). Substrate was classified as fines or sand only when it fully buried underlying substrate (i.e., a fine layer of silt overlying a substrate of larger size was classified as the larger particle). Percent substrate cover in each riffle was estimated as the percentage of 100 measured particles in each substrate class.
Size and abundance of interstitial refuges was also measured within each of the four quadrats using short lengths of clear plastic tubing inserted into substrate interstices, as described by Finstad et al. (2007). Tubing with diameters of 0.7, 1, 1.9, and 2.2 cm (Fig. S2) were used to probe and count interstices with a minimum length of 3 cm or greater, measured using the largest diameter tube that would fit within each potential refuge. The area of each identified interstitial refuge was then estimated by multiplying the diameter of the refuge by its length.
Fish and benthic invertebrate collection
Fish were collected from all riffles (i.e., control and treatment riffles), pre- and post-restoration by triple-pass, depletion electrofishing in August of 2021 and 2022, respectively (2021 and 2022 Scientific Fish Collection Permits SU21-624949 and SU22-718288). To prevent the escape of fish from the sample area, fish-tight stop nets were carefully installed across the wetted width of the channel at the upstream and downstream ends of riffles before fish collection. Capture effort (i.e., time spent electroshocking) was held constant for each pass in a single riffle. A conservative estimate of fish abundance in each riffle was calculated as the sum of the fish caught in all three passes, rather than a depletion estimate because low or inconsistent catches in Pepin Creek riffles precluded meaningful depletion estimates; density was estimated as total abundance divided by the surface area of the riffle. Capture efficiency of Nooksack Dace by electrofishing is in the range of 25%–35% (Bonamis 2011; Avery-Gomm et al. 2014), so that estimates of density from electrofishing could be rescaled to approximate true densities by multiplying by a factor of 3.3. However, rescaling Nooksack Dace density estimates would not have changed the relative treatment effects and so abundance estimates were left unexpanded. A minimum of 10 min separated the end of one electrofishing pass and the commencement of the next to allow the recovery of fish that escaped collection. Collected fish were sedated with MS2-22, identified, and measured for weight, and total length. Fish were placed in flow-through bins to recover prior to their release at the site of capture. Sampling procedures involving the collection of Endangered Nooksack Dace were designed and implemented in accordance with applicable laws (Fisheries and Oceans Canada Species at Risk Permits: 21-HPAC-00744 and 21-HPAC-00746) and approved by the Animal Care Committee at the University of British Columbia (permit: A21-0086) and followed Canadian Council for Animal Care guidelines.
Samples of benthic invertebrates were also collected from all replicate riffles in Pepin and Bertrand Creeks before and after restoration. A 15 cm wide Surber sampler (0.023 m2 sampling area) with 250 μm mesh was used to collect four benthic samples from each riffle in late July of 2021 and 2022, approximately two weeks prior to fish collection. Each sample was collected from a randomly chosen point at either 25%, 50%, or 75% across each of four evenly spaced transects across each riffle.
Samples were collected by disturbing the substrate by hand to a depth of 5 cm to dislodge benthic invertebrates into the downstream Surber sampler. Each sample was filtered through a 250 μm mesh sieve and preserved in 70% ethanol. Invertebrates were subsequently sorted from debris in the laboratory at 16 times magnification using a stereomicroscope and identified to family or subfamily according to Merritt and Cummins (1996). The length of each invertebrate was measured by digitizing images projected from the microscope onto an adjacent digitizing pad using a drawing tube (Roff and Hopcroft 1986). The dry weight biomass of each taxa was then estimated using published length–weight regressions for aquatic and terrestrial invertebrates (Benke et al. 1999; Sabo et al. 2002). To estimate the available biomass of prey for—dace, benthic invertebrate taxa were considered potential prey if they were previously documented in the diet of Nooksack Dace or considered vulnerable to predation, after Avery-Gomm et al. (2014) and McPhail (1996).
Data analysis
We performed two general analyses to understand the response of Nooksack Dace to both natural and experimental variation in substrate composition. First, we evaluated the relationships between substrate variables and dace density using univariate or multiple regression. Second, we assessed the response of both habitat and dace density to riffle restoration as a BACI experiment, testing for an interaction between time (before or after restoration) and treatment as the diagnostic for a significant restoration effect. All analyses were completed using R version 4.1.1 (R Core Team 2022).
Using the regression approach, the influence of substrate attributes on dace density (dace·m−2) across all riffles combined were evaluated as fixed effects in separate generalized linear mixed models (GLMM). To meet model assumptions, a Tweedie distribution with a log link function was used (Tweedie package; Dunn and Smyth 2005). Stream system (Bertrand Creek vs. Pepin Creek) and restoration site (i.e., treatment replicates) were included as nested, random effects. The effects of substrate attributes on Nooksack Dace density were evaluated as separate univariate GLMM due to small sample size where the inclusion of additional predictor variables could reduce the statistical power of the model. Model fit was assessed using Q–Q plots of (DHARMa package; Hartig 2022) and by verifying deviations from the uniformity of randomized quantile residuals (Kolmogorov–Smirnov test; performance package; Lüdecke et al. 2021).
The density of interstitial refuges within a riffle was calculated as the number of identified refuges in a quadrat divided by quadrat area (0.16 m2). Area (two-dimensional footprint) of individual interstitial refuges was calculated as the product of average refuge length and diameter for each of the four diameter classes. The response of average and maximum dace total length to average interstitial refuge area (all diameter classes combined) was modelled using separate linear mixed models (LMM), with stream system and restoration site included as nested, random effects. Residuals were tested for normality and homogeneity using Shapiro–Wilks and Breusch–Pagan tests, respectively (R Core Team 2022). The effect of cobble and gravel substrate cover on the density and area of interstitial refuges was also modelled using separate LMM. To meet model assumptions of homogeneity, response variables were log transformed when modeling the effects on cobble and gravel substrate cover on the density of interstitial refuges. The effect of sand substrate cover on the average density and area of interstitial refuges was modelled using separate GLMM, as described previously.
In addition to direct modelling of habitat and substrate effects on dace density as described above, we applied also analyzed the BACI design using a LMM to evaluate the effects of the riffle restoration intervention on substrate composition, interstitial refuge area and density, and Nooksack Dace density (Roni et al. 2018). The diagnostic for a significant treatment effect was an interaction between time (two levels: before and after restoration) and restoration treatment (cobble, gravel, or control), with time and treatment as fixed effects and stream and restoration site treated as nested, random effects in the LMM. To normalize model residuals, response variables were square-root transformed for all LMMs with the exception of interstitial refugia area, which remained untransformed, and sand and gravel substrate cover, where the dependent variables were log transformed. Normality and homogeneity of residuals were tested using Shapiro–Wilk and Breusch–Pagan tests, respectively (R Core Team 2022). Lastly, Wilcoxon Rank Sum tests were used to separately analyze changes in overall dace density and cobble substrate cover in Bertrand Creek and Pepin Creek. Wilcoxon Rank Sum tests were used because transformations could not normalize residuals.
The effects of substrate composition and interstitial refuges on benthic invertebrate biomass were assessed using separate LMM, as described above, with log transformations of invertebrate biomass to normalize residuals. To test for a positive effect of invertebrate biomass on dace density, GLMM was used, with stream system and restoration site treated as nested, random effects. A Wilcoxon Rank Sum test was used to compare invertebrate biomass between the two study streams. Lastly, a BACI design was used to examine whether riffle restoration had an effect on invertebrate biomass using LMM, as described above.
Results
Response of Nooksack Dace to substrate composition and abundance of interstitial refuges
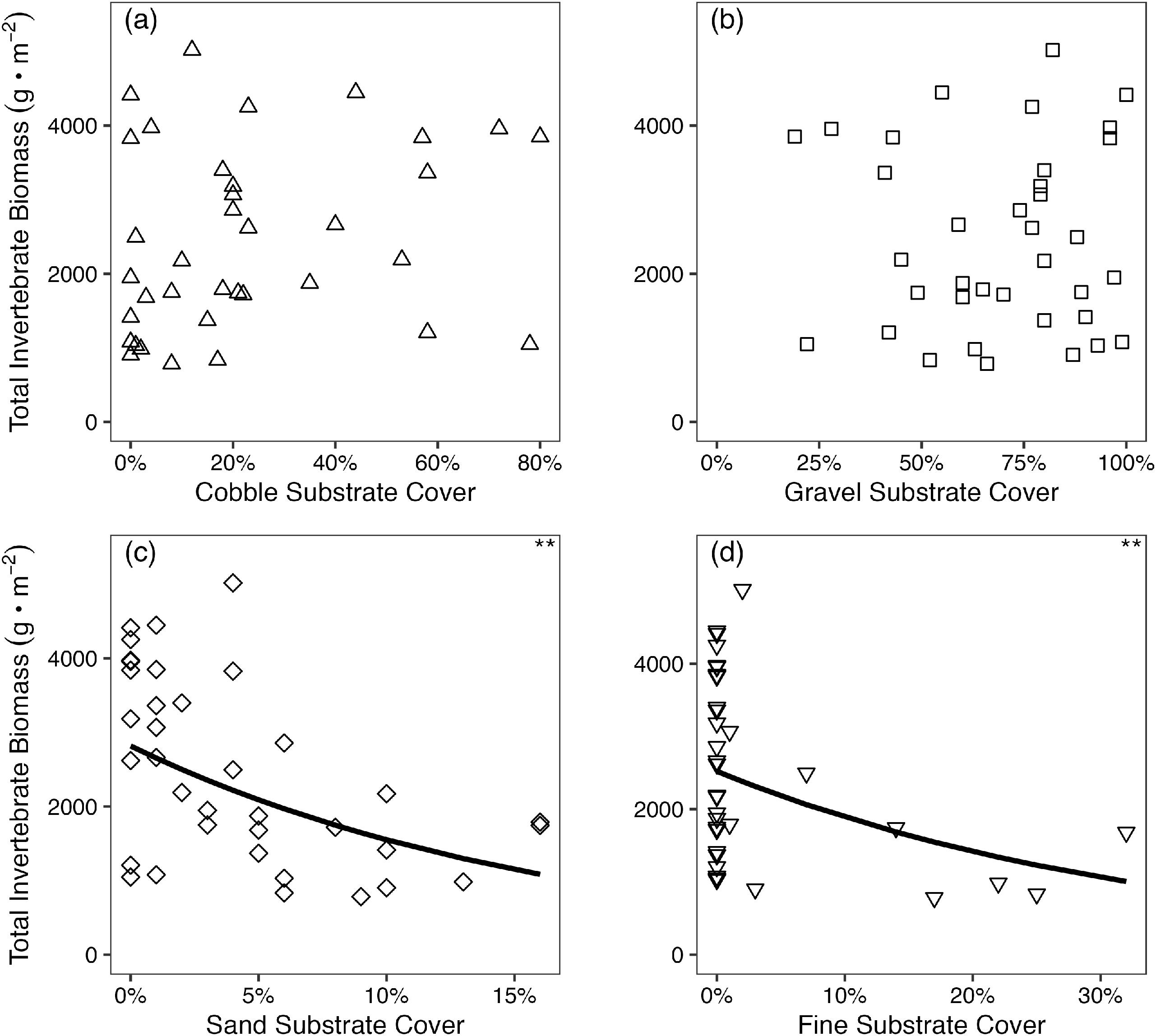
As expected, dace density increased with cobble substrate cover (Fig. 2a; AIC = 62.2, p < 0.001; Table 1) and conversely declined with increasing gravel and sand substrate cover (Figs. 2b and 2c; AIC = 70.9, p < 0.001 for gravel; AIC = 87.1, p = <0.001 for sand; Table 1). Dace density also had a strong negative association with fine substrate cover, with an average density of 0.83 dace/m2 when fine substrate cover was absent, and 0.17 dace/m2 when fine substrate exceeded 0% cover (Fig. S2).
Fig. 2.
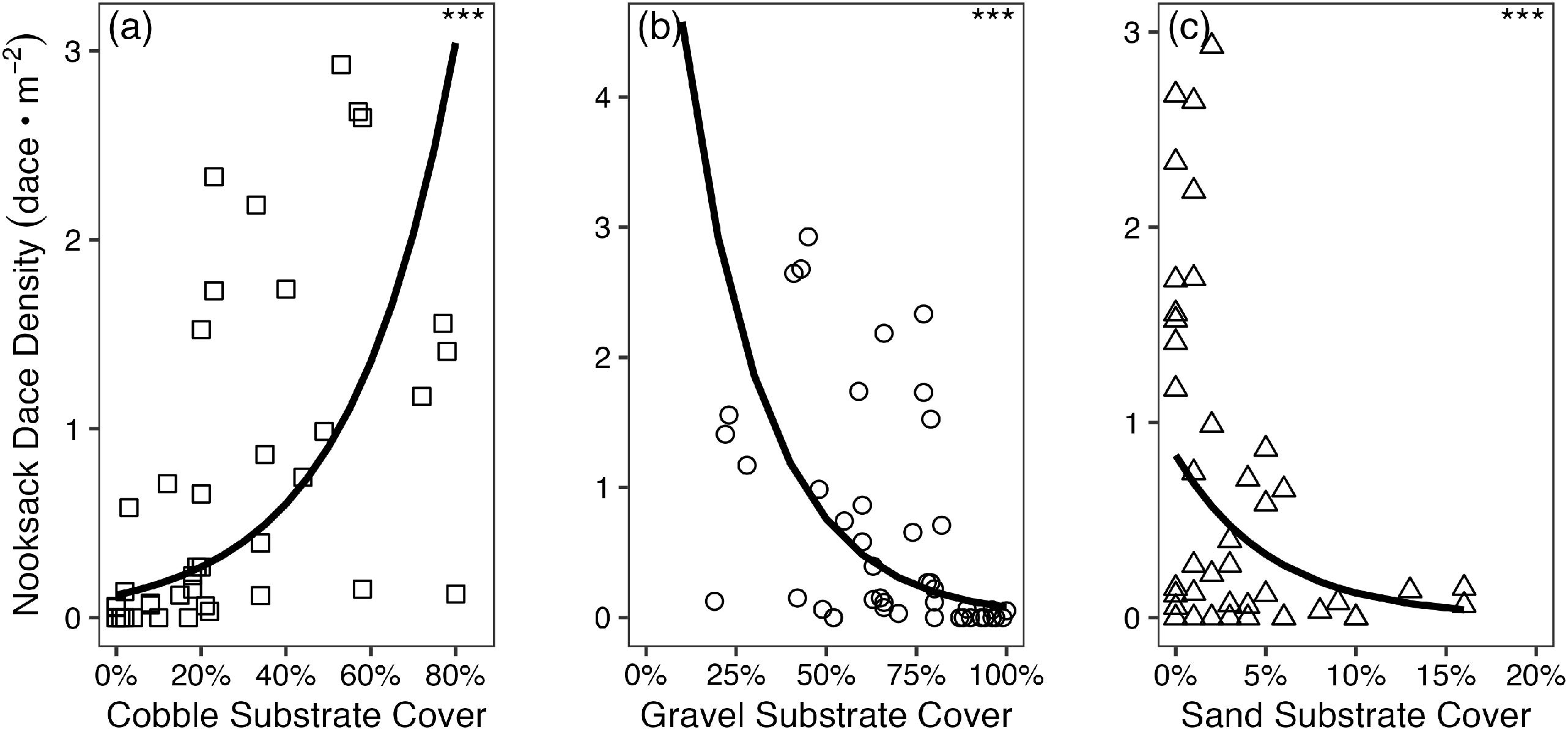
Table 1.
| Variables | ||||||||
|---|---|---|---|---|---|---|---|---|
| Response (yi) | Predictor (xi) | System | Model | Equationa | AIC | p value | n | Transformations |
| Dace density (y1) | Cobble cover (%; x1) | Bertrand Creek | GLMM | 60.9 | <0.001 | 44 | – | |
| Cobble cover (%; x1) | Pepin Creek | GLMM | y1 = 10 (0.04x1 − 2.02) | 60.9 | <0.001 | 44 | – | |
| Gravel cover (%; x2) | Bertrand Creek | GLMM | 70.0 | <0.001 | 44 | – | ||
| Gravel cover (%; x2) | Pepin Creek | GLMM | y1 = 10 (− 0.04x2 + 1.87) | 70.0 | <0.001 | 44 | – | |
| Sand cover (%; x3) | Bertrand Creek | GLMM | 85.6 | <0.001 | 43 | – | ||
| Sand cover (%; x3) | Pepin Creek | GLMM | y1 = 10 (− 1.19x3 − 0.07) | 85.6 | <0.001 | 43 | – | |
| Refugia density (x4) | Bertrand Creek | GLMM | 69.5 | <0.001 | 44 | – | ||
| Refugia density (x4) | Pepin Creek | GLMM | y1 = 10 (0.04x4 − 1.73) | 69.5 | <0.001 | 44 | – | |
| Refugia area (x5) | Bertrand Creek | GLMM | 75.1 | <0.001 | 44 | – | ||
| Refugia area (x5) | Pepin Creek | GLMM | y1 = 10 (0.30x5 − 2.25) | 75.1 | <0.001 | 44 | – | |
| Refugia density (y2) | Cobble cover (%; x1) | Bertrand Creek | LMM | 132.6 | <0.001 | 44 | log | |
| Cobble cover (%; x1) | Pepin Creek | LMM | y2 = 10 (0.04x1 − 1.41) | 132.6 | <0.001 | 44 | log | |
| Gravel cover (%; x2) | Bertrand Creek | LMM | 144.9 | <0.001 | 44 | log | ||
| Gravel cover (%; x2) | Pepin Creek | LMM | y2 = 10 (− 0.04x2 + 5.06) | 144.9 | <0.001 | 44 | log | |
| Sand cover (%; x3) | Bertrand Creek | GLMM | y2 = 10 (− 0.12x3 + 3.35) | 329.8 | <0.001 | 43 | – | |
| Sand cover (%; x3) | Pepin Creek | GLMM | y2 = 10 (− 0.12x3 + 3.35) | 329.8 | <0.001 | 43 | – | |
| Refugia area (y3) | Cobble cover (%; x1) | Bertrand Creek | LMM | y3 = (0.09x1 + 1.84) | 186.2 | <0.001 | 44 | – |
| Cobble cover (%; x1) | Pepin Creek | LMM | y3 = (0.09x1 + 1.84) | 186.2 | <0.001 | 44 | – | |
| Gravel cover (%; x2) | Bertrand Creek | LMM | y3 = (− 0.09x2 + 10.43 + 0.20) | 194.4 | <0.001 | 44 | – | |
| Gravel cover (%; x2) | Pepin Creek | LMM | y3 = (− 0.09x2 + 10.43) | 194.4 | <0.001 | 44 | – | |
| Sand cover (%; x3) | Bertrand Creek | GLMM | y3 = 10 (− 0.08x3 + 1.64) | 195.3 | <0.001 | 43 | – | |
| Sand cover (%; x3) | Pepin Creek | GLMM | y3 = 10 (− 0.08x3 + 1.64) | 195.3 | <0.001 | 43 | – | |
a
Mixed-effect models incorporated system (Bertrand Creek vs. Pepin Creek) as a random effect.
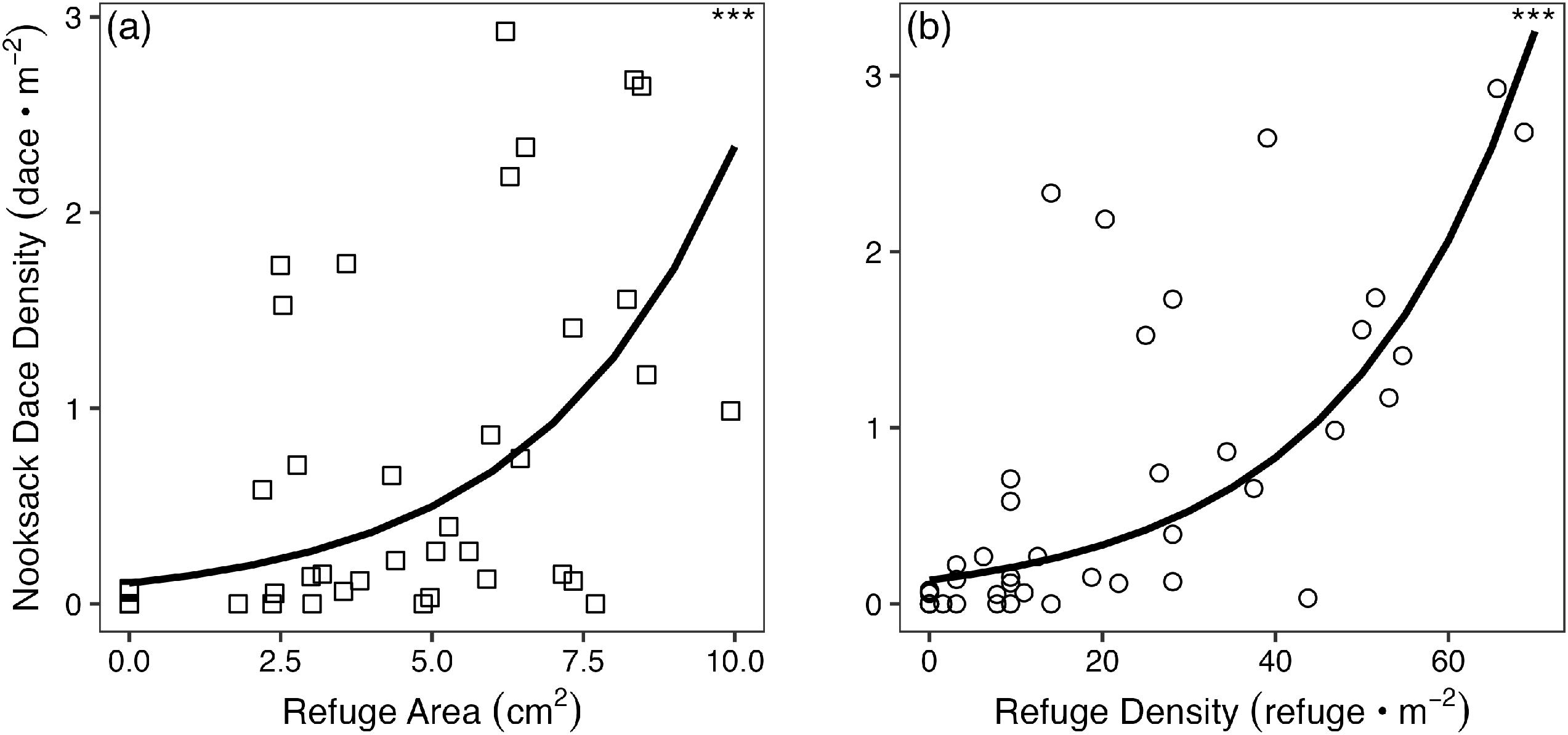
Dace density also increased with average interstitial refugia area (Fig. 3a; AIC = 77.1, p = <0.001; Table 1) and density (Fig. 3b; AIC = 68.9, p = <0.001; Table 1), supporting the inference that a positive association of dace with large substrate is driven, in part, by the availability of interstitial habitat.
Fig. 3.
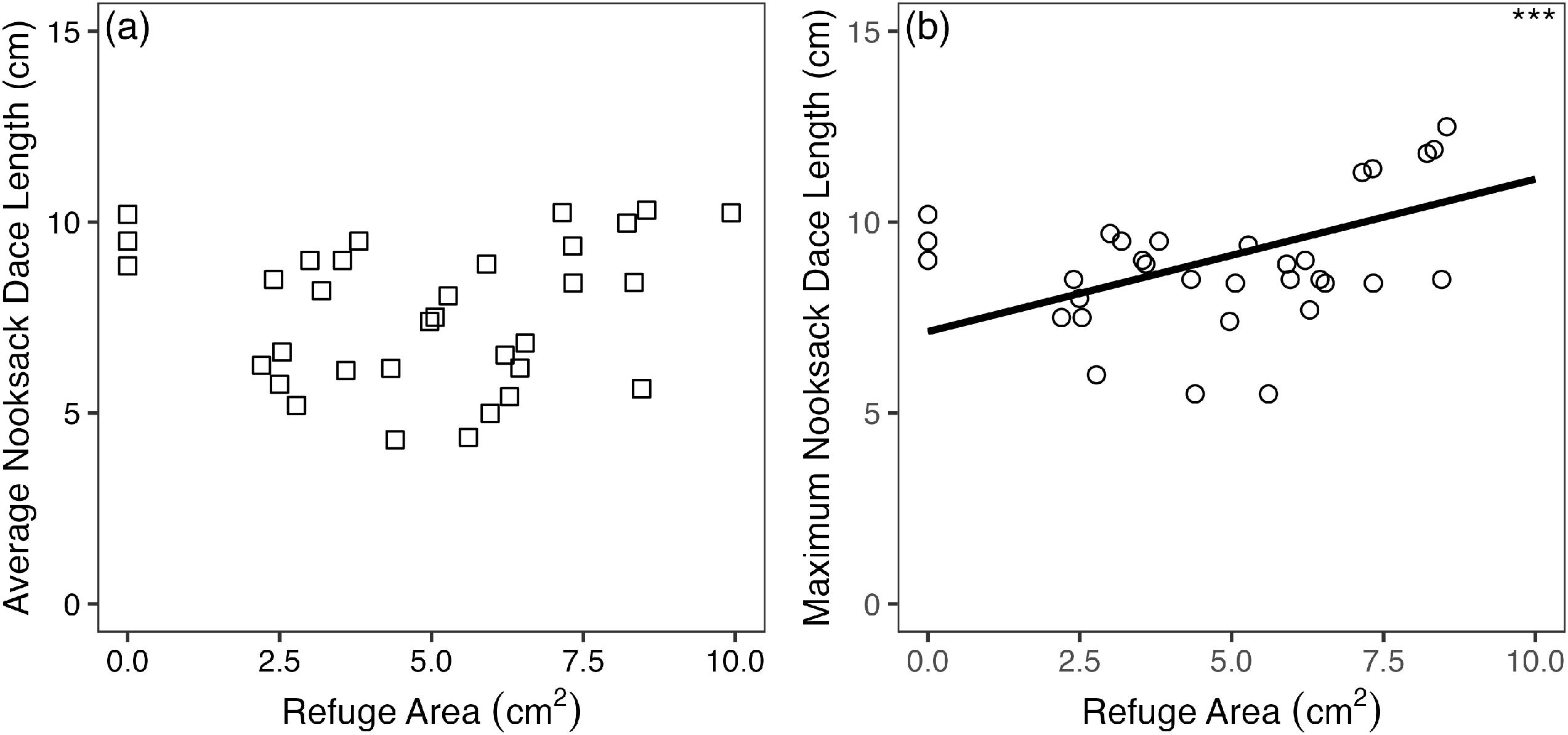
Average length of dace collected in a riffle appeared unrelated to average interstitial refugia area (Fig. 4a; AIC = 107.9, p = 0.2) however, the maximum length of dace collected in a riffle increased with average interstitial refugia area (Fig. 4b; AIC = 136.8, p < 0.001).
Fig. 4.
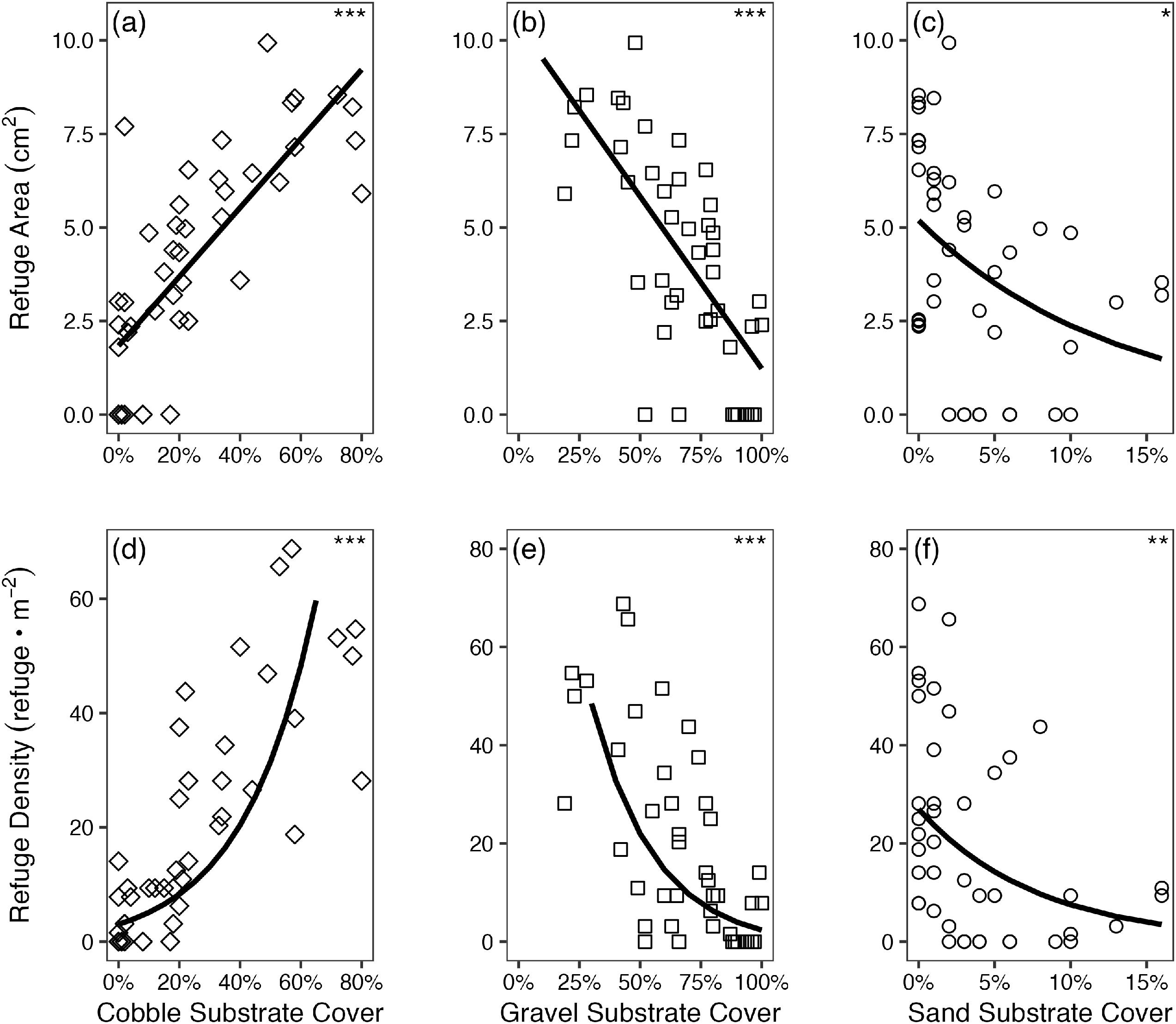
As expected, both the area and density of interstitial refuges was positively correlated with cobble substrate cover (Figs. 5a and 5d; p < 0.001; p < 0.001, respectively; Table 1). Conversely, interstitial refugia area and density declined with increasing cover of gravel and sand (Figs. 5b, 5c, 5e, and 5f; p < 0.001; p < 0.001; p = 0.04; p = 0.002; Table 1; note that one outlier data point had to be removed to improve normality of the regression of refuge area and density on sand in panel 5c and 5f).
Fig. 5.
Effect of riffle substrate compositions on benthic invertebrate biomass
Substrate composition varied considerably among riffles but appeared to have limited effect on total benthic invertebrate biomass (Fig. 6); however, total benthic invertebrate biomass did decrease with increasing percentage sand and fine substrate cover (Figs. 6c and 6d; p = 0.001 and 0.007, respectively). Similarly, the subset of total benthic biomass that was classified as prey for Nooksack Dace was unrelated to either average interstitial refuge area (linear regression, F1,35 = 1.5, R2 = 0.04, p = 0.2) or density (linear regression, F1,35 = 0.8, R2 = 0.03, p = 0.4).
Fig. 6.
Invertebrate prey biomass was highly variable, and was not a significant predictor of dace density (Fig. S5; GLMM, AIC = 86.6; p = 0.6), although average total invertebrate prey biomass (W = 232 and p = 0.03) was higher in Bertrand Creek (2193 mg·m−2) then in Pepin Creek (1640 mg·m−2).
Effectiveness of riffle habitat restoration
Addition of cobbles resulted in a significant increase in Nooksack Dace density, unlike gravel or control treatments, which was reflected in a significant time by treatment interaction (Fig. 7a; p = <0.001). The average area and density of interstitial refuges increased across all treatments in response to habitat restoration but was only significant in cobble treatments (Figs. 7b and 7c; p = 0.003 and 0.03, respectively). However, while there was a significant increase in the average area and density of interstitial refuges within cobble treatments, there were no significant time by treatment interactions for interstitial refuge area and density (p = 0.1 and 0.2, respectively), likely because interstitial refuge density showed an increasing trend in all substrate treatments post-restoration, including in controls.
Fig. 7.
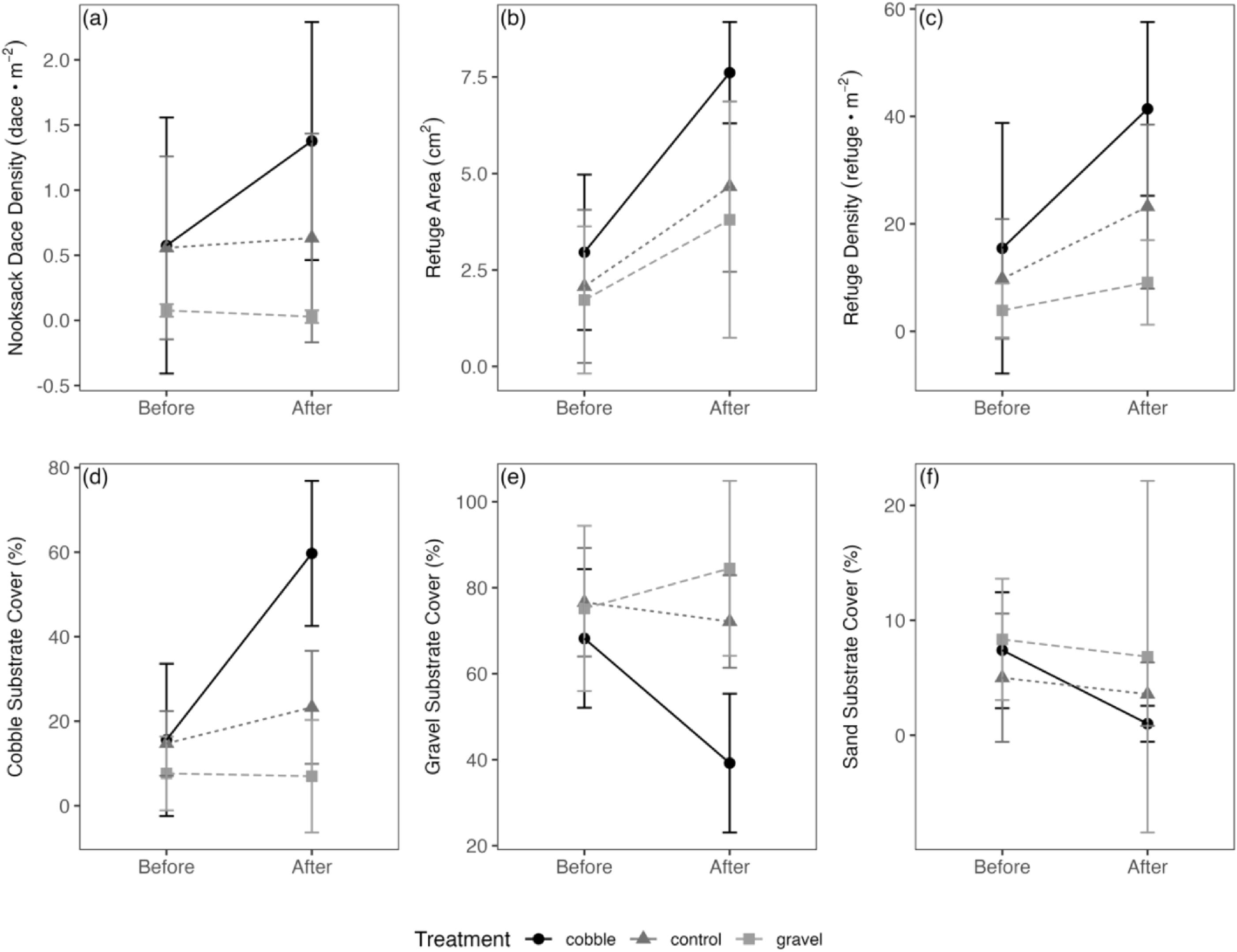
As expected, there was a significant interaction between substrate cover and time, with cobble substrate cover increasing significantly in the cobble treatments (p = 0.01) but not in gravel or control treatments, validating the effectiveness of the habitat manipulations on substrate composition (Fig. 7d). Similarly, gravel substrate cover declined following cobble addition (Fig. 7e; p = 0.01) and sand declined following both cobble and gravel addition (Fig. f; p = 0.003 and 0.01, respectively) but not in control treatments. Contrary to expectation, total benthic invertebrate prey biomass did not increase in response to restoration (Fig. S5; no significant time by treatment interaction, p = 0.9), although power to detect a treatment effect was very low due to high variation.
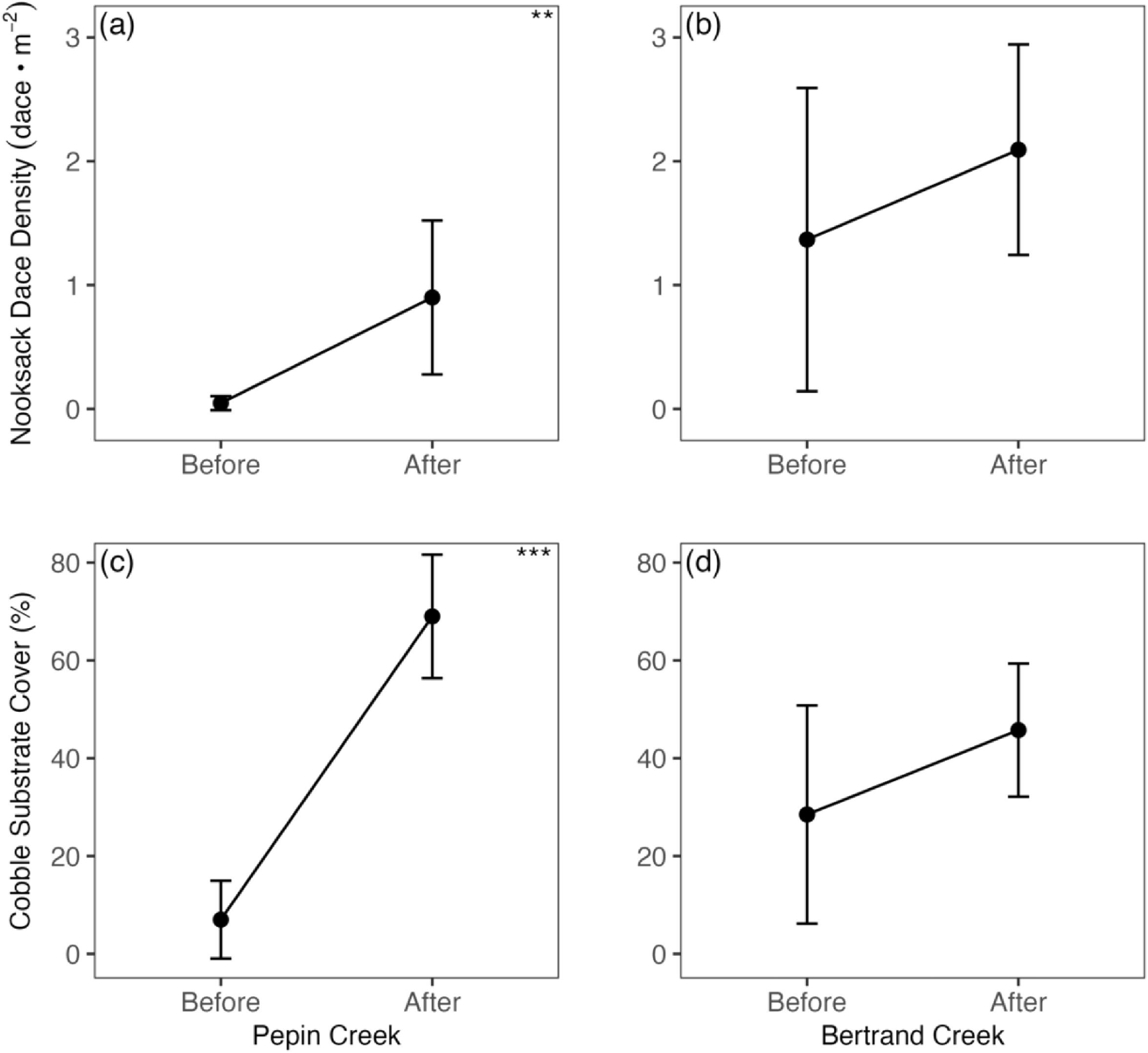
The effect of cobble addition on dace density was significantly different between Pepin and Bertrand Creeks (Figs. 8a and 8b), which differed in their initial cobble cover, with coarse substrate initially scarce in Pepin Creek but relatively abundant in Bertrand Creek pre-restoration (Figs. 8c and 8d). Consequently, we anticipated a large response to cobble addition in Pepin Creek, and minimal to no response in Bertrand Creek. However, while the relative increase in both dace density and cobble cover was much higher in Pepin Creek (W = 35, p = 0.008 and W = 36, p = 0.002, respectively), the absolute increase in dace densities were similar in both streams. This suggests that cobble remained limiting to dace in Bertrand Creek, despite a much higher initial proportion of cobble substrate, although the increase in dace density was only statistically significant in Pepin Creek.
Fig. 8.
Discussion
The restoration of degraded habitat has become widespread in freshwaters and is a key element in the recovery of many species at risk (Tear et al. 1993; Follstad Shah et al. 2007; Reid et al. 2016; Gawecka and Bascompte 2021). However, the effectiveness of habitat restoration is rarely assessed (Bernhardt and Palmer 2011), despite evaluation being an essential step in adaptive management. In this study, we examined whether Nooksack Dace density increased in response to riffle habitat restoration to assess whether it represents an effective recovery strategy for this Endangered species. We explicitly tested two general hypotheses for the potential role of coarse cobble substrate in supporting higher dace abundance: (i) that substrate interstices would provide increased habitat for benthic invertebrates, which increases prey abundance and therefore densities of benthic insectivores like Nooksack Dace and (ii) that interstices directly provide physical habitat for dace.
Benthic invertebrate data do not support the inference of enhanced dace abundance from increased benthic invertebrate prey biomass in cobble substrate, or a correlation between invertebrate prey biomass and interstitial space. This suggests that the primary mechanism whereby cobble affects Nooksack Dace is through direct use of interstitial habitat, rather than increased prey abundance. This is supported by significant correlations between the area and density of interstitial space and the maximum size and density of dace, respectively. However, this conclusion must be tempered by the large variation in benthic invertebrate biomass in all substrate treatments (Fig. S5), and the correspondingly low power to detect differences in benthic invertebrate biomass between gravel and cobble treatments, and controls. Many studies have demonstrated higher benthic invertebrate biomass on coarser substrate (e.g., Burdon et al. 2013; Scholl et al. 2022), and there was a detectable negative correlation between benthic biomass and areal coverage of fines and sand in this study (Fig. 6). Both Bertrand Creek and Pepin Creek are mildly eutrophic streams with high algal production, which tends to generate high benthic macroinvertebrate abundance (Moore and Palmer 2005; Bernot et al. 2006; Herringshaw et al. 2011). Generally high benthic invertebrate biomass may mask any relationships between interstitial space and invertebrate biomass that might manifest in a more oligotrophic system.
Overall, this study confirmed that Nooksack Dace rely on unembedded interstitial spaces in coarse substrate, presumably for refuge from flow and predation, for spawning, and for foraging (Pearson et al. 2007; McEwan and Joy 2014; Kawanishi et al. 2015; Champion et al. 2018). The observation of reduced dace abundance when these interstitial spaces are lost is broadly consistent with the effects of sediment deposition seen elsewhere (Wood and Armitage 1997; Rambaud et al. 2009; Descloux et al. 2013). Unexpectedly, the riffle restoration experiment also coincided with record fall floods that took place in November 2021 after the initial dace baseline sampling, but before the terminal sampling in August 2022. Bertrand Creek received peak flows approaching 50 m3·s−1 (unpublished data), raising concerns over potential Nooksack Dace mortality associated with bedload movement that could confound the experimental design. However, there was no reduction in dace density following the floods, despite evidence of significant bedload movement, indicating that Nooksack Dace are well-adapted to coping with high discharge events and bed instability. The increase in interstitial refuge area and density in control substrate treatments in the second year suggests that the flood exported fine sediment and reduced overall embeddedness. While this effect could confound the experimental treatment to some extent, the lack of a dace response in gravel and control treatments suggests that the overall reduction in fines was not consequential for the outcome of the study.
An increase in fish density following habitat restoration can be caused by one (or all) of three mechanisms. First, creation of higher quality habitat can lead to immigration of fish from adjacent lower quality habitat (e.g., dace may abandon riffles with highly embedded substrate and move to newly created riffles with abundant interstitial space; Roni 2019). This aggregation response does not necessarily demonstrate an increase in total population size, despite the local increase in fish density in restored habitat, because it could simply represent a redistribution of fish. However, net immigration to the newly created habitat and higher fish densities relative to pre-restoration supports the inference of higher quality habitat at the restored sites, assuming that fish follow an ideal free distribution (i.e., that density reflects habitat quality; Kennedy and Gray 1993). If the fish population in question is not recruitment limited (i.e., there are adequate juvenile fish to saturate adult habitat; Rosenfeld and Hatfield 2006), then it is reasonable to assume that any lower quality habitat vacated by immigrants would be filled by recruits, resulting in a net increase in population size and carrying capacity. In contrast, if the population is severely recruitment limited, then increasing adult habitat may not result in a population increase, even if fish relocate to restored habitat.
Second, increased growth and survival in higher quality restored riffles could cause increased fish density (i.e., through lower mortality) and increase overall population size (Roni 2019), even without immigration of fish to restored habitat. However, any population-level effect through this mechanism would likely be smaller than for a population with abundant recruitment to saturate both new and existing habitat. Finally, if reproduction takes place in the restored riffle habitat (as might be expected with Nooksack Dace), then enhanced recruitment would complement increased adult density or growth, and maximize the likelihood of a rapid population response to restoration. Young-of-the-year Nooksack Dace were present in all sampled riffles in Bertrand Creek, indicating that in situ recruitment may have contributed to increased densities in restored riffles (although young-of-the-year fish could also have originated from unrestored riffles). In contrast, while Pepin Creek riffles clearly showed a strong increase in dace density, no young-of-the-year recruits were observed in any of the restored (or control) riffles over two years of sampling in Pepin Creek (Gray 2023). This indicates that the higher post-restoration density in Pepin Creek was entirely an aggregation/immigration response, and is unlikely to reflect a net increase in population size given the extreme recruitment limitation that we observed. This serves as a cautionary note that increased density following habitat restoration may not always be an accurate diagnostic of successful population recovery, and highlights the importance of considering the mechanism(s) underlying locally increased density. Low and intermittent recruitment is clearly limiting the Nooksack Dace population in Pepin Creek, and simple riffle restoration failed to alleviate this, at least for one year of post-restoration monitoring. Loss of juvenile rearing habitat due to encroaching reed canarygrass is implicated in juvenile recruitment failure in Pepin Creek (Gray and Rosenfeld 2024), where riffle restoration will remain ineffective in population recovery until the juvenile recruitment bottleneck is addressed.
This study integrated a structured BACI habitat restoration experiment with subsidiary analyses to identify the mechanisms driving abundance of Endangered Nooksack Dace, and to evaluate the practical effectiveness of riffle restoration. While replicates were limited by logistic challenges (i.e., resources, time, and space) as is often the case for field studies, the strong and consistent treatment effects suggest that the results are robust and biologically relevant. The distribution of Nooksack Dace in Canada is limited to only four streams, with the majority of fish residing in the two study streams, so that the observed dynamics are directly relevant to over half of the total Nooksack Dace population in Canada. The reaches where experiments were conducted were also highly representative of general habitat conditions in each study stream. And by encompassing the full range of habitat quality from very high dace densities to strongly recruitment limited, the experimental design ensured a broad scope of inference and generality across a wide range of habitat contexts.
Restoring degraded habitat represents one potential technique to reverse the effects of anthropogenic disturbance by recovering habitat that has been degraded, damaged or destroyed (Rey Benayas et al. 2009). Significant increases in Nooksack Dace density following riffle restoration indicates that riffle restoration does indeed represent a useful tool in the recovery of an endangered riffle specialist, but the overall role of this intervention in species recovery needs to be kept in perspective, for two reasons. First, riffle restoration will only be effective at increasing population size in streams where (i) recruitment is sufficient to saturate restored habitat and (ii) in streams where any net gain in habitat through restoration is not negated by an equivalent habitat loss elsewhere (Maron et al. 2018). The management paradigm of no net loss of habitat (Harper and Quigley 2005) is based on the reasonable inference that populations will decline if habitat loss exceeds habitat gain. The ultimate effectiveness of local habitat restoration in recovering populations is predicated on the assumption that habitat loss elsewhere is negligible. Habitat restoration to compensate for ongoing habitat destruction elsewhere is generally not a sustainable or effective routine management strategy (Harper and Quigley 2005; Moreno-Mateos et al. 2015) because the rate of habitat loss generally exceeds capacity to restore degraded habitat (Maron et al. 2012, 2018). Second, instream habitat restoration, even when locally effective, takes place in a watershed context (Beechie et al. 2010) where cumulative basin-scale impacts can reverse local gains. Of particular concern in Nooksack Dace watersheds are increases in water temperature and hypoxia associated with eutrophication from cumulative landscape-scale nutrient enrichment and climate change, which can easily render restored reaches uninhabitable for the target species (Rosenfeld et al. 2021). To be effective, instream habitat restoration must be combined with watershed-scale management to ensure that restored habitat remains functional, and is part of a larger integrated recovery strategy. In the absence of wider habitat protections, restored habitat may eventually be degraded or destroyed, as has occurred with riffle habitat throughout the LFV (Boyle et al. 1997; Pearson et al. 2007).
References
Aadland L.P. 1993. Stream habitat types: their fish assemblages and relationship to flow. North American Journal of Fisheries Management, 13(4): 790–806.
Arthington A.H., Dulvy N.K., Gladstone W., Winfield I.J. 2016. Fish conservation in freshwater and marine realms: status, threats and management. Aquatic Conservation: Marine and Freshwater Ecosystems, 26: 838–857.
Avery-Gomm S., Rosenfeld J.S., Richardson J.S., Pearson M. 2014. Hydrological drought and the role of refugia in an endangered riffle-dwelling fish, Nooksack dace (Rhinichthys cataractae sp.). Canadian Journal of Fisheries and Aquatic Sciences, 71: 1625–1634.
Beechie T.J., Sear D.A., Olden J.D., Pess G.R., Buffington J.M., Moir H., et al. 2010. Process-based principles for restoring river ecosystems. Bioscience, 60(3): 209–222.
Benke A.C., Huryn A.D., Smock H.L., Bruce J. 1999. Length-mass relationships for freshwater macroinvertebrates in North America with particular reference to the southeastern United States. Journal of the North American Benthological Society, 18(3): 308–343.
Berg M.A., Allen D.M. 2007. Low flow variability in groundwater-fed streams. Canadian Water Resources Journal, 32(3): 227–246.
Bernhardt E.S., Palmer M.A. 2011. River restoration: the fuzzy logic of repairing reaches to reverse catchment scale degradation. Ecological Applications, 21(6): 1926–1931.
Bernot M.J., Tank J.L., Royer T.V., David M.B. 2006. Nutrient uptake in stream draining agricultural catchments of the Midwestern United States. Freshwater biology, 51(3): 499–509.
Bo T., Fenoglio S., Malacarne G., Pessino M., Sgariboldi F. 2007. Effects of clogging on stream macroinvertebrates: an experimental approach. Limnologica, 37(2): 186–192.
Bonamis A. 2011. Utilization of two-stage single-pass electrofishing to estimate abundance and develop recovery-monitoring protocols for the endangered Nooksack dace (Rhinichthys cataractae) in Canada. Simon Fraser University, Burnaby, British Columbia, Canada. 67pp.
Boyd L., Grant P., Lemieux J., Iacarella J.C. 2022. Cumulative effects of threats on at-risk species habitat in the Fraser Valley, British Columbia. Canadian Manuscript Report of Fisheries and Aquatic Sciences, 3243: viii + 65.
Boyle C.A., Lavkulich L., Schreier H., Kiss E. 1997. Changes in land cover and subsequent effects on Lower Fraser Basin ecosystems from 1827 to 1990. Environmental Management, 21: 185–196.
Burdon F.J., McIntosh A.R., Harding J.S. 2013. Habitat loss drives threshold response of benthic invertebrate communities to deposited sediment in agricultural streams. Ecological Applications, 23(5): 1036–1047.
Champion J.M., Rosenfeld J.S., Shadwick R. 2018. Effects of water velocity and substrate composition on foraging efficiency of an endangered benthic cyprinid, Nooksack dace (Rhinichthys cataractae subsp. cataractae). Hydrobiologia, 805: 231–243.
Clapcott J.E., Young R.G., Harding J.S., Matthaei C.D., Quinn J.M., Death R.G. 2011. Sediment Assessment Methods: Protocols and guidelines for assessing the effects of deposited fine sediment on in-stream values. Cawthron Institute, Nelson, New Zealand.
Collen B., Whitton F., Dyer E., Baillie J., Cumberlidge N., Darwall W., et al. 2014. Global patterns of freshwater species diversity, threat and endemism. Global Ecology and Biogeography, 23: 40–51.
COSEWIC. 2007. COSEWIC assessment and update status report on the Nooksack dace Rhinichthys cataractae ssp. in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. pp. vii + 27.
COSEWIC. 2018. COSEWIC assessment and status report on the Nooksack Dace Rhinichthys cataractae in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. xii + 38pp. Available from http://www.registrelep-sararegistry.gc.ca/default.asp?lang=en&n=24F7211B-1 [accessed 25 July 2023].
de Graaf I., Gleeson T., van Beek L., Sutanudjaja E., Bierkens M. 2019. Environmental flow limits to global groundwater pumping. Nature, 574: 90–94.
Descloux S., Datry T., Marmonier P. 2013. Benthic and hyporheic invertebrate assemblages along a gradient of increasing streambed colmation by fine sediment. Aquatic Sciences, 75(4): 493–507.
Dudgeon D., Arthington H.A., Gessner O.M., Kawabata Z., Knowler J.D., Lévêque C., et al. 2006. Freshwater biodiversity: importance, threats, status and conservation challenges. Biological Reviews, 81(2): 163–182.
Duncan J., Lockwood J. 2001. Extinction in a field of bullets: a search for causes in the decline of the world's freshwater fishes. Biological Conservation, 102(1): 97–105.
Dunn P.K., Smyth G.K. 2005. Series evaluation of Tweedie exponential dispersion model densities. Statistics and Computing, 15: 267–280.
Finstad A.G., Einum S., Forseth T., Ugedal O. 2007. Shelter availability affects behaviour, size-dependent and mean growth of juvenile Atlantic salmon. Freshwater Biology, 52: 1710–1718.
Fisheries and Oceans Canada. 1998. Wild, threatened, endangered and lost streams of the lower Fraser Valley summary report: lower Fraser Valley stream review vol. 3. Fraser River Action Plan, Habitat and Enhancement Branch, Fisheries and Oceans Canada, Vancouver.
Fisheries and Oceans Canada. 2020. Recovery strategy for the Nooksack Dace (Rhinichthys cataractae ssp.) in Canada. First amendment. Species at Risk Act Recovery Strategy Series. Fisheries and Oceans Canada, Ottawa. vii+ 47pp.
Florsheim J.L., Mount J.F., Chin A. 2008. Bank erosion as a desirable attribute of rivers. Bioscience, 58(6): 519–529.
Follstad Shah J., Dahm C.N., Gloss S.P., Bernhardt E.S. 2007. River and riparian restoration in the southwest: results of the national river restoration science synthesis project. Restoration Ecology, 15(3): 550–562.
Gawecka K.A., Bascompte J. 2021. Habitat restoration in spatially explicit metacommunity models. Journal of Animal Ecology, 90: 1239–1251.
Göthe E., Wiberg-Larsen P., Kristensen E.A., Baattrup-Pedersen A., Sandin L., Friberg N. 2015. Impacts of habitat degradation and stream spatial location on biodiversity in a disturbed riverine landscape. Biodiversity and Conservation, 24(6): 1423–1441.
Gray J. 2023. Evaluation of habitat features responsible for variations in the abundance of endangered Nooksack dace (Rhinichthys cataractae sp. cataractae) and the role of habitat restoration in their recovery. MSc. thesis, University of British Columbia, Vancouver, British Columbia, Canada.
Gray J., Rosenfeld J.S. 2024. Habitat effects on recruitment, population limitations, and recovery potential in an endangered stream fish. Canadian Journal of Fisheries and Aquatic Sciences, Just-IN.
Green R.H. 1979. Sampling Design and Statistical Methods for Environmental Biologists. Wiley, Chichester, UK.
Harper D.J., Quigley J.T. 2005. No net loss of fish habitat: a review and analysis of habitat compensation in Canada. Environmental Management, 36(3): 343–355.
Hartig F. 2022. DHARMA: Residuals Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. Available from http://florianhartig.github.io/DHARMa/ [accessed 22 April 2024].
Herringshaw C.J., Stewart T.W., Thompson J.R., Anderson P.F. 2011. Land use, stream habitat and benthic invertebrate assemblages in a highly altered Iowa watershed. The American Midland Naturalist, 165(2): 274–293.
Jelks H.L., Walsh S.J., Burkhead N.M., Contreras-Balderas S., Diaz-Pardo E., Hendrickson D.A., et al. 2008. Conservation status of imperiled North American freshwater and diadromous fishes. Fisheries, 33(8): 372–407.
Kawanishi R., Dohi R., Fujii A., Inoue M. 2015. Effects of sedimentation on an endangered benthic fish, Cobitis shikokuensis: is sediment-free habitat a requirement or a preference? Ecology of Freshwater Fish, 24(4): 584–590.
Kemp P., Sear D., Collins A., Naden P., Jones I. 2011. The impacts of fine sediment on riverine fish. Hydrological Processes, 25(11): 1800–1821.
Kennedy M., Gray R.D. 1993. Can ecological theory predict the distribution of foraging animals? A critical analysis of experiments on the ideal free distribution. Oikos, 68(10): 158–166.
Lennox A.P. III, Rasmussen B.J. 2016. Long-term effects of channelization on a cold-water stream community. Canadian Journal of Fisheries and Aquatic Sciences, 73: 1530–1537.
Lüdecke D., Ben-Shachar M., Patil I., Waggoner P., Makowski D. 2021. “performance: an R package for assessment, comparison and testing of statistical models”. Journal of Open Source Software, 6(60): 3139.
Maron M., Brownlie S., Bull J.W., Evans M.C., von Hase A., Quétier F., et al. 2018. The many meanings of no net loss in environmental policy. Nature Sustainability, 1: 19–27.
Maron M., Hobbs R.J., Moilanen A., Matthews J.W., Christie K., Gardner T.A., et al. 2012. Faustian bargains? Restoration realities in the context of biodiversity offset policies. Biological Conservation, 155: 141–148.
McEwan A.J., Joy M.K. 2014. Diel habitat use of two sympatric galaxiid fishes (Galaxias brevipinnis and G. postvectis) at two spatial scales in a small upland stream in Manawatu, New Zealand. Environmental Biology of Fishes, 97: 897–907.
McPhail J.D. 1996. COSEWIC status report on Nooksack dace Rhinichthys sp. in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. 1–8pp.
Merritt R.W., Cummins K.W. 1996. An Introduction to the Aquatic Insects of North America. Kendall/Hunt Publishing Company, Dubuque. 862p.
Miranda R., Miqueleiz I., Farwall W., Sayer C., Dulvy N., Carpenter K., et al. 2022. Monitoring extinction risk and threats of the world's fishes based on the Sampled Red List Index. Reviews in Fish Biology and Fisheries, 32: 975–991.
Moore A.A., Palmer M.A. 2005. Invertebrate biodiversity in agricultural and urban headwater streams: implications for conservation and management. Ecological Applications, 15(4): 1169–1177.
Moreno-Mateos D., Maris V., Béchet A., Burran M. 2015. The true loss caused by biodiversity offsets. Biological Conservation, 192: 552–559.
Naden P.S., Murphy J.F., Old G.H., Newman J., Scarlett P., Harman M., et al. 2016. Understanding the controls on deposited fine sediment in the streams of agricultural catchments. Science of the Total Environment, 547: 366–381.
Pearson M.P. 1998. Habitat Inventory and Enhancement Needs for the Endangered Salish Sucker (Catostomus Sp.) and Nooksack Dace (Rhinichthys Sp.). Fisheries Project Report No. 76.
Pearson M.P. 1999. The biology and management of the Salish Sucker and Nooksack Dace. Proceedings of a Conference on the Biology and Management of Species and Habitats at Risk, Kamloops, BC. pp. 15–19.
Pearson M.P. 2004. The ecology, status, and recovery potential of Nooksack dace (Rhinichthys cataractae ssp.) and Salish sucker (Catostomus sp.) in Canada. Ph.D. thesis, University of British Columbia, Vancouver, Canada.
Pearson M.P., Hatfield T., McPhail J.D., Richardson J.S., Rosenfeld J.S., Schreier H., et al. 2007. Recovery strategy for the Nooksack Dace (Rhinichthys cataractae) in Canada. Species at Risk Act Recovery Strategy Series, Fisheries and Oceans Canada, Vancouver. pp.vi + 31.
Poff N.L., Huryn A.D. 1998. Multi-scale determinants of secondary production in Atlantic salmon (Salmo salar) streams. Canadian Journal of Fisheries and Aquatic Sciences, 55: 201–217.
R Core Team. 2022. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Available from https://www.R-project.org/ [accessed 27 September 2022].
Rabeni C., Smale M. 1995. Effects of siltation on stream fishes and the potential mitigating role of the buffering riparian zone. Hydrobiologia, 303: 211–219.
Rambaud M., Combroux I., Haury J., Moret J., Machon N., Zavodna M., Pavoine S. 2009. Relationships between channelization structures, environmental characteristics, and plant communities in four French streams in the Seine–Normandy catchment. Journal of the North American Benthological Society, 28(3): 596–610.
Reid A., Carlson A., Creed I., Eliason E., Gell P., Johnson P., et al. 2019. Emerging threats and persistent conservation challenges for freshwater biodiversity. Biological Reviews, 94: 849–873.
Reid B.N., Thiel R.P., Peery M.Z. 2016. Population dynamics of endangered blanding's turtles in a restored area. The Journal of Wildlife Management, 80(3): 553–562.
Rey Benayas J.M., Newton A.C., Diaz A., Bullock J.M. 2009. Enhancement of biodiversity and ecosystem services by ecological restoration: a meta-analysis. Science, 325(5944): 1121–1124.
Reynolds J., Webb T., Hawkins L. 2005. Life history and ecological correlates of extinction risk in Europeans freshwater fishes. Canadian Journal of Fisheries and Aquatic Sciences, 62(4): 854–862.
Roff J.C., Hopcroft R.R. 1986. High precision microcomputer based measuring system for ecological research. Canadian Journal of Fisheries and Aquatic Sciences, 43(10): 2044–2048.
Roni P. 2019. Does river restoration increase fish abundance and survival or concentrate fish? The effects of project scale, location, and fish life history. Fisheries, 44: 7–19.
Roni P., Anders P.A., Beechie T.J., Kaplowe D.J. 2018. Review of tools for identifying, planning, and implementing habitat restoration for Pacific Salmon and Steelhead. North American Journal of Fisheries Management, 38: 355–376.
Rosenfeld J., Pearson M.P., Miners J., Zinn K. 2021. Effects of landscape-scale hypoxia on Salish Sucker and salmonid habitat associations: implications for endangered species recovery and management. Canadian Journal of Fisheries and Aquatic Sciences, 78(9): 1219–1233.
Rosenfeld J.S., Hatfield T. 2006. Information needs for assessing critical habitat of freshwater fish. Canadian Journal of Fisheries and Aquatic Sciences, 63(3): 683–698.
Sabo J.L., Bastow J.L., Power M.E. 2002. Length-mass relationships for adult aquatic and terrestrial invertebrates in a California watershed. Journal of the North American Benthological Society, 21(2): 336–343.
Scholl E., Cross W., Guy C. 2022. Geomorphology shapes relationships between animal communities and ecosystem function in large rivers. Oikos, 2023(2): e09431.
Suttle K., Power M., Levine J., McNeely C. 2004. How fine sediment in riverbeds impairs growth and survival of juvenile salmonids. Ecological Applications, 14(4): 969–974.
Taylor E.B., McPhail J.D., Ruskey J.A. 2015. Phylogeography of the longnose dace (Rhinichthys cataractae) species group in northwestern North America—the Nooksack dace problem. Canadian Journal of Zoology, 93: 727–734.
Tear T.H., Scott J.M., Hayward P.H., Griffith B. 1993. Status and prospects for success of the endangered species act: a look at recovery plans. Science, 262(5136): 976–977.
U.S. Geological Survey. 2023. National Water Dashboard at https://dashboard.waterdata.usgs.gov/app/nwd/en/?aoi=default [accessed 10 April 2023].
van der Lee A., Koops M. 2015. Are small fishes more sensitive to habitat loss? A generic size-based model. Canadian Journal of Fisheries and Aquatic Sciences, 73: 716–726.
Vélez-Espino L.A., Koops M. 2012. Capacity for increase, compensatory reserves, and catastrophes as determinants of minimum viable population in freshwater fishes. Ecological Modelling, 247: 319–326.
Walters D., Leigh D., Bearden A. 2003. Urbanization, sedimentation, and the homogenization of fish assemblages in the Etowah River Basin, USA. Hydrobiologia, 494: 5–10.
Wentworth K.C. 1922. A scale of grade and class terms for clastic sediments. The Journal of Geology, 30: 377–392.
Wolman M.G. 1954. A method of sampling coarse river bed material. Eos, Transactions American Geophysical Union, 35(6): 951–956.
Wood P.J., Armitage P.D. 1997. Biological effects of fine sediment in the lotic environment. Environmental Management, 21(2): 203–217.
Supplementary material
Supplementary Material 1 (PDF / 484 KB).
- Download
- 484.22 KB
Information & Authors
Information
Published In

FACETS
Volume 9 • Number 1 • January 2024
Pages: 1 - 15
Editor: Trevor Pitcher
History
Received: 26 November 2023
Accepted: 17 May 2024
Version of record online: 29 August 2024
Notes
This paper is part of a collection entitled Progress and Priorities for the Recovery of Aquatic Species at Risk in Canada.
Copyright
© 2024 The Author(s). This work is licensed under a Creative Commons Attribution 4.0 International License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Data Availability Statement
Data generated or analyzed during this study are available from the corresponding author or at the online repository Figshare.
Key Words
Sections
Subjects
Authors
Author Contributions
Conceptualization: JG, JR, MP
Data curation: JG, JR, KC, JR
Formal analysis: JG, JR
Funding acquisition: JR
Investigation: JG, JR, MP, KC, JR
Methodology: JG, JR, MP
Project administration: JG, JR
Resources: JG, JR, MP
Software: JG
Supervision: JG, JR, MP
Validation: JG, JR, MP
Visualization: JG, JR, MP
Writing – original draft: JG
Writing – review & editing: JG, JR, MP, KC, JR
Competing Interests
The authors declare there are no competing interests.
Funding Information
Funding: This research was supported by the federal Department of Fisheries and Oceans Pacific Region MOU 15 to the British Columbia Ministry of Environment.
Metrics & Citations
Metrics
Other Metrics
Citations
Cite As
John Gray, Jordan Rosenfeld, Mike Pearson, Kay Colletti, and Jeremy Ross. 2024. The effect of riffle restoration on the recovery of endangered Nooksack Dace (Rhinichthys cataractae sp. cataractae). FACETS.
9: 1-15.
https://doi.org/10.1139/facets-2023-0217
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
There are no citations for this item