Investigating the potential uptake of microplastic-derived carbon into a boreal lake food web using carbon-13 labelled plastic
Abstract
Microplastics degrade slowly over time, leaching carbon (C) that could be subsequently incorporated into aquatic food webs. Current estimates of microplastic degradation vary, and little is known about microplastic-derived C fate under natural environmental conditions. To investigate whether microplastics leach C that is subsequently incorporated into aquatic food webs, we added isotopically enriched microplastics to Lake 378 at the Experimental Lakes Area in Ontario, Canada. In an ∼1100 L limnocorral (in situ open-bottom enclosure), we added 99% 13C-labelled polystyrene (8–216 µm in longest dimension) at a nominal concentration of 3268 particles/L. A second limnocorral without microplastics served as a negative control. Monthly measurements of δ13C-DIC and δ13C-DOC in filtered water revealed no detectable leaching of 13C from the plastic. Compound-specific isotope analysis of δ13C in amino acids of bulk plankton and periphyton revealed a slight (0.5‰) enrichment in 13C, within the range of natural variability for these organisms. Under the natural conditions of temperate oligotrophic lakes, degradation of microplastics is likely a very slow process that was not possible to detect in this 4-month experiment. Future studies should focus on assessing degradation of microplastics under realistic field scenarios to improve estimates of degradation pathways and associated time scales.
Introduction
There is growing concern about the fate and effects of microplastics (plastics < 5 mm in size) in aquatic ecosystems due to their abundance and persistence in the environment, and their potential toxic effects on aquatic organisms (Horton et al. 2017; Wang et al. 2019; Bucci et al. 2020). Plastic polymers are synthesized primarily from fossil fuels like petroleum or natural gases, and thus are carbon (C)-based products. Due to the large amount of plastic entering aquatic ecosystems, plastic-derived C could be considered a significant C pool in the environment (Dees et al. 2021).
Slowly, macroplastics are fragmented through abiotic degradation (i.e., photo- and mechanical degradation) in surface water, leading to the generation of microplastics, which are more susceptible to further biodegradation by microbes (Chamas et al. 2020; Zhu et al. 2020; Dees et al. 2021; Priya et al. 2022). During these processes, degradation of microplastics can result in the release (termed “leaching”) of small dissolved organic carbon (DOC) compounds from microplastics into water. Further, microbial utilization of microplastics can lead to mineralization to CO2 and CH4 (Curran and Strlič 2015; Zhu et al. 2020; Lee et al. 2021; Priya et al. 2022). In addition to the fragmentation and mineralization of macro and microplastics in the environment, microplastics can foster microbial growth more than natural DOC, suggesting that microplastics could increase microbial activity and enhance the mineralization of microplastics in the environment (Sheridan et al. 2022).
Previously, compound-specific isotope analysis (CSIA) revealed that in laboratory experiments, microbial degradation of microplastics resulted in mineralization to respired CO2 and incorporation of plastic C in cell membrane components of microbes (Taipale et al. 2019, 2023). Further, this plastic-derived C was then trophically transferred and utilized by algae and zooplankton (Taipale et al. 2019, 2023). These studies suggest that MP-derived C can be effectively used by mixotrophic algae in the formation of biomolecules. However, the degradation and subsequent incorporation of plastic-derived C into aquatic food webs has not been studied in the natural environment and a better understanding of these processes under such conditions is needed (Sander et al. 2019).
The objective of this experiment was to measure the leaching of C from microplastics over time in a boreal lake ecosystem and determine if we could detect the leached C within the aquatic food web. We employed isotopically labelled 13C-polystyrene (13C-PS) as a novel approach to trace MP-derived C in an aquatic food web (Liu et al. 2024). We added 13C-PS to a 1100 L limnocorral (in-lake enclosure, open to sediment and enclosing natural water and biota of the lake environment). Potential plastic leaching and incorporation into the food web was measured over time using stable isotope analysis of δ13C-DIC (dissolved inorganic carbon), δ13C-DOC, and δ13C in amino acids (δ13CAA) of periphyton and plankton. We hypothesized that (1) microplastics in a lake lose C over time due to both abiotic (photo-oxidative) and biotic (microbial) degradation, and (2) that DOC and DIC derived from microplastics is bioavailable and incorporated into the food web through utilization by microbes. From these hypotheses, we predicted that (1) DIC and DOC would be 13C-enriched (relative to a control; measured as δ13C-DIC and δ13C-DOC) following addition of 13C labelled microplastics, and (2) amino acids in plankton and periphyton would be 13C-enriched (relative to a control; measured as δ13C in amino acids) following addition of 13C labelled microplastics.
Materials and methods
Experimental design and set-up
This experiment took place in two limnocorrals (i.e., in-lake enclosures) in a natural boreal lake at the International Institute for Sustainable Development Experimental Lakes Area (ELA; Ontario, Canada). Our study lake, Lake 378 (49°42′38″N, 93°46′32″W), is an oligotrophic temperate lake with a surface area of 251 579 m2 and maximum depth of 16.6 m and has low background levels of microplastics (McIlwraith et al. 2024). This study is one of a series of experiments being conducted under the “pELAstic Project” over several years to understand the fate and effects of microplastics in lake ecosystems (https://thepelasticproject.com/). Because this is the first time, to our knowledge, that 13C-plastic was intentionally added to a natural aquatic ecosystem, the study was intended to be exploratory in nature.
In May 2021, two 1 m wide open-bottomed limnocorrals were set up in the littoral zone at a depth of 1 m in ELA Lake 378 (Supporting Information, Part A: Fig. S1). The limnocorrals were constructed with a polyvinylchloride floatation ring attached to a food-grade polyethylene curtain (Curry Industries, Winnipeg, Manitoba, Canada) that was sealed to the sediment using a double ring of sandbags. The food-grade polyethylene curtain was used to limit the potential for plastic C to leach from the curtains into the limnocorral water. These limnocorrals were open to the lake sediments and the atmosphere so that they enclosed the natural sediment, water, and biota from the lake. The food-grade polyethylene “walls” of the limnocorrals were impermeable such that there was no exchange between water in the lake and the limnocorrals once sealed to the lake bottom. The estimated volume of each limnocorral was 1100 L. A tracer was not used in the present study to monitor leakage from the limnocorral or exchange of water between the limnocorral and the lake through sediment. However, based on previous studies using the same design and set up techniques, leakage through these mechanisms was anticipated to be negligible and we do not expect diffusion through sediment to be a relevant route of exchange that would significantly dilute our 13C-labelled plastic signature in the current study (Orihel et al. 2006; Graves et al. 2021).
For this experiment, we used 99% 13C-labelled PS that was synthesized and ground to a powder of microplastic fragments (Supporting Information, Part A: Fig. S2) by MilliporeSigma (Burlington, Massachusetts, USA). We used carbon-13 labelled PS because it allows us to clearly differentiate between plastic-derived C and natural C that is composed of 98.9% carbon-12. Prior to addition, we counted and characterized the 13C-PS fragments in the laboratory. Briefly, the size range and concentration of 13C-PS was characterized by first transferring the weighed and ground 13C-PS into a clean 1 L nalgene bottle with 100 mL of multi-Q ultrapure water. Two small scoops of Sparkleen™ low foam lab detergent were added to prevent clumping of microplastics. A 0.5 mL aliquot of 13C-PS solution was pipetted onto a 0.2 um polycarbonate filter paper. Particles were counted under a microscope at 80x magnification with an eye piece at 10x magnification. The subsampling and counting procedure was done for a total of five replicates, and the counts per replicate were <10% different from the mean. Following counting, photographs of the field of view with microplastics were taken for each replicate and later loaded in ImageJ to measure the length of each particle. Since the fragments were irregularly shaped, measurements were standardized by measuring only the longest side of each particle. Over 200 particles were measured across the five replicates counted and particles ranged in size from 8 to 216 µm (mean: 53.4 µm; median: 50.9 µm) (Supporting Information, Part A: Fig. S3).
To one limnocorral, we added 50 mg (3268 000 particles or 3268 particles/L) of 13C-PS. The 13C-PS was first mixed into 1 L of lake water in a high-density polyethylene (HDPE) Nalgene container in the laboratory. This solution was then added to the mesocosm by uncapping the 1 L bottle underwater and gently shaking the bottle, held horizontally, to release the plastic into the mesocosm. The bottle was rinsed with mesocosm water and the process repeated twice more. The other limnocorral served as a control where no plastic was added. To ensure that enough 13C-labelled PS was being added to the limnocorral to detect degradation of microplastics, prior to beginning the field experiment, we used background C information from the lake and previously calculated rates of microplastic degradation from a previous laboratory study by Zhu et al. (2020) to estimate the projected rate of C leaching from plastic. From this, we estimated the anticipated enrichment of 13C in water column dissolved C (see calculations in Supporting Information, Part B). For these calculations, we assumed that microplastics would mix approximately evenly within the water column of the 1 m wide by 2 m deep limnocorral. Polystyrene is a neutrally buoyant microplastic and we assumed, based on this feature, that most PS would remain in the water column. Elagami et al. (2022) estimated the residence time of PS under typical lake conditions and found that particle size had great influence on water residence time. Based on their models, our range of 13C-PS particles was expected to remain in the water column up to 10 000 days (Elagami et al. 2022). As such, our sampling efforts focused on collecting water and biota from this compartment within the limnocorrals.
Sample collection and processing
Since the degradation of microplastic is a very slow process (Taipale et al. 2019, 2022, and 2023), we aimed to conduct a long-term (4 month) study to detect the leaching of C from microplastics under natural conditions. The frequency and timing of sampling collection were based on previous work by Taipale et al. (2019) showing that the mineralization of microplastic is a slow process and that in a closed system, the total amount of plastic-derived C, and associated changes in δ13C, should increase over time, such that conducting a long experiment is necessary to detect C leaching from plastic. Sampling times throughout the experimental period are referred to as weeks pre- or post-additions, with week 0 being the week of the 13C-PS addition. Starting on 17 May (week −1, e.g., 1 week prior to additions), one integrated water sample was taken from the middle of the water column (0.5 m) in each limnocorral using a peristaltic pump for δ13C analysis of DOC, DIC, and for water chemistry. These water samples were collected on weeks −1, 1, 4, 8, 12, and 17 of the experimental period. For δ13C-DOC and δ13C-DIC, an inline filter with stacked 0.7 µm GF/F and 0.2 µm polycarbonate filters were used to minimize biological activity, and to exclude microplastics from water samples. Water (60 mL) was collected into acid-cleaned HDPE bottles with no head space and transported to the laboratory on ice. Water samples were immediately preserved in ZnCl2 (0.6 mL of 50% w/v) and stored at 4 °C until further analysis. Additional water samples were collected to measure DIC, DOC, alkalinity, and conductivity. For these samples, water was again collected from 0.5 m depth in the limnocorral using a peristaltic pump. For DIC, water was collected into a 150 mL HDPE septum cap bottle with no headspace. For DOC, samples were collected in a 1 L HDPE bottle and transported to the laboratory where they were filtered through 0.7 µm GF/F filter. The DIC and DOC were analyzed in-house following Stainton et al. (1977). Waters for alkalinity and conductivity were collected into 150 mL septum cap HDPE bottles and measured in-house at the IISD-ELA.
In addition to water, one composite sample of plankton and one of periphyton were collected for isotopic analyses at the end of the experiment (i.e., week 17) so that biota had the entire 4 month experimental period to incorporate plastic-derived C. We collected bulk plankton by towing horizontally in each mesocosm with a 53 µm mesh tow net. We also suspended strips (made of the same polyethylene material as the walls; 10 cm wide × 1 m deep) vertically in each limnocorral to collect periphyton, which were subsequently recovered at the end of the experiment. Biota samples were flash-frozen in liquid nitrogen and then stored at −80 °C until further processing for CSIA. To investigate enrichment of 13C in biota as evidence of MP-derived C incorporation, we employed amino acid CSIA rather than bulk isotope analysis because organisms could inevitably contain 13C labelled microplastics. Bulk isotope analysis would not be able to differentiate microplastic particles and from biota tissue and could inflate our 13C value.
Experiment quality assurance/quality control measures
To minimize contamination of the 13C-labelled PS in the control limnocorral during sampling, limnocorrals were always sampled in the order of control followed by treatment. Separate equipment including different sets of tubing, filter holders, forceps were used for each limnocorral and were kept in separate storage containers. Gloves were worn at all times during sampling and a clean hands/dirty hands approach was used to minimize contamination of equipment with water from the 13C-PS limnocorral.
Compound-specific isotope analysis
Carbon stable isotope ratios in DIC and DOC were measured at the Environmental Isotope Laboratory at University of Waterloo (UW-EIL) using continuous flow-isotope ratio mass spectrometry (CF-IRMS) according to methods described by St. Jean (2003) and Stainton et al. (1977). Carbon stable isotopes are reported as parts per thousand (per mille or ‰) relative to Vienna PeeDee Belemnite and are expressed using the notation δ13C. Quality assurance/quality control measures included duplicate runs of every fourth sample with an average percent difference (±standard deviation) of 1.6% (±1.8%) and an analytical accuracy of ±0.2‰.
Carbon stable isotopes of amino acids (δ13CAA) were analyzed at the Dalhousie University Stable Isotope Biogeochemistry Laboratory using methods from Silfer et al. (1991) and modified in Chen et al. (2022). Briefly, ∼5 mg aliquots of the plankton and periphyton samples were hydrolyzed in 2 mL of 6 N HCl at 110 °C for 20 h. Hydrolysates were spiked with a norleucine (Nle) internal standard and purified using cation exchange chromatography (Dowex 50WX8 200-400 H). Amino acids were converted to isopropyl-trifluoroacetyl derivatives and further purified using solvent phase extraction (chloroform and buffered phosphate). Samples were injected in triplicate on a Thermo Trace 1310 gas chromatograph coupled via GC Isolink II and Conflo IV interfaces to a Thermo Delta V isotope ratio mass spectrometer. Sample injections were bracketed by triplicate injections of an external standard consisting of a synthetic mix of 13 amino acids prepared in the same way as the sample hydrolysates. Measured δ13CAA values were corrected for instrument drift and linearity effects and calibrated to the external amino acid standard using equations in Silfer et al. (1991) to correct for additional C atoms introduced during derivatization. Analytical reproducibility, as measured from the standard deviation of sample triplicate injections, averaged 0.5‰ across all amino acids. As a check on data accuracy, the calibrated δ13C values of Nle, as well as all 13 amino acids in laboratory running standards (chlorella powder and fish muscle) co-prepared with the samples, averaged within 0.5‰ of known values.
Statistical analyses
The R software environment (version 4.3.2; Posit team, 2024) was used for all statistical analyses and alpha was set a 0.05 for all statistical tests. Normal distribution of data was confirmed using Shapiro–Wilks normality tests. We highlight that the present study was designed to explore the use of 13C-labelled microplastic to investigate fate under realistic environmental conditions and that the interpretation of our results is limited given the lack of replication in this experiment. As such, we focused on the enrichment of 13C in the treatment limnocorral as an indicator that MP-derived C was present in DIC, DOC, or amino acids. Analysis of covariance was used to determine if there were differences in δ13C-DIC or δ13C-DOC over time between the control and 13C-PS limnocorrals. For amino acids, one-tailed, paired t tests were used to determine whether treatment values were significantly 13C enriched relative to controls. Further, the available literature was used to determine the ranges of values measured for these samples in boreal lakes to estimate ranges of natural variability for these endpoints (Supporting Information, Part A: Tables S1 and S2; Bade et al. 2004; Zigah et al. 2011; Larsen et al. 2013). Enrichment of 13C beyond natural variability in DOC, DIC, and amino acids was used as an additional indicator of MP-derived C incorporation. Following the completion of our experiment, we used a more recently published degradation rate (Vesamaki et al. 2022) to re-calculate the projected rate of C leaching from plastic in our limnocorral (see calculations in Supporting Information, Part C) to obtain a second projected rate for comparison to our field study. Finally, we estimated the degradation rate of microplastics in the current study using the relative difference in δ13C-DOC in the control versus treatment limnocorral after 4 months (see calculations in Supporting Information, Part D).
Results
Physicochemical conditions in limnocorrals
Physicochemical parameters varied over the experimental period and the conditions were similar across the control and treatment limnocorrals. Temperature in the water column of the limnocorrals ranged from 13.7 to 28.0 °C in the control and 13.9 to 28.1 °C in the 13C-PS treatment over the experimental period (Supporting Information, Part A: Fig. S4). Dissolved inorganic carbon ranged from 127 to 211 µmol/L in the control and 129–182 µmol/L in the treatment limnocorral over time (Table 1; Supporting Information, Part A: Fig. S5). DIC increased over time in both the control and treatment limnocorrals; although DIC appeared to increase more rapidly in the control, the pattern was only marginally nonsignificant (test for homogeneity of slopes, F3,8 = 6.41, p = 0.06, R2 = 0.52, Table 1; Supporting Information, Part A: Fig. S5). DOC ranged from 606 to 762 µmol/L in the control and 593 to 978 µmol/L in the treatment limnocorral (test for homogeneity of slopes, F3,8 = 33.2, p = 0.005, R2 = 0.87, Table 1; Supporting Information, Part A: Fig. S5); DOC increased over time in the control and treatment limnocorrals, but the increase was greater in the treatment limnocorral (Table 1). Alkalinity also increased over time in both limnocorrals (Table 1; Supporting Information, Part A: Fig. S5).
Table 1.
| Limnocorral | Week | δ13C-DIC (‰) | δ13C-DOC (‰) | DIC (µmol/L) | DOC (µmol/L) | Particulate C (µg/L) | Particulate N (µg/L) | Alkalinity (µEq/L) | Conductivity (µS/cm) |
|---|---|---|---|---|---|---|---|---|---|
| Control | −1 | −10.2 | −27.5 | 127 | 606 | 761 | 97 | 138 | − |
| Control | 1 | −9.5 | −26.9 | 139 | 622 | 832 | 122 | 134 | 21 |
| Control | 4 | −8.2 | −28.1 | 170 | 611 | 1259 | 128 | 189 | 27 |
| Control | 8 | −13.40 | −26.9 | 159 | 729 | 489 | 48 | 203 | 25.5 |
| Control | 12 | −11.1 | −28.1 | 217 | 632 | 512 | 54 | 217 | 25.5 |
| Control | 17 | −10.4 | −27.4 | 211 | 762 | 579 | 59 | 197 | 23.5 |
| 13C-PS | −1 | −10.2 | −26.5 | 129 | 609 | 724 | 95 | 143 | − |
| 13C-PS | 1 | −10.5 | −26.2 | 150 | 593 | 736 | 107 | 152 | 22 |
| 13C-PS | 4 | −10.2 | −29.4 | 161 | 638 | 851 | 111 | 218 | 24.5 |
| 13C-PS | 8 | −15.5 | −26.8 | 139 | 764 | 562 | 67 | 225 | 30.5 |
| 13C-PS | 12 | −10.7 | −26.0 | 157 | 750 | 1876 | 158 | 238 | 28.8 |
| 13C-PS | 17 | −12.5 | −26.3 | 182 | 978 | 442 | 34 | 209 | 24.4 |
δ13C-DIC and DOC in limnocorrals over time
Carbon stable isotope ratios of DIC ranged from –13.4‰ to –8.2‰ in the control and –15.5‰ to –10.2‰ in the treatment limnocorral over the 17-week experiment (Table 1). Similarly, δ13C-DOC values changed little over the experimental period, ranging from –28.1‰ to –26.9‰ in the control and –29.3‰ to –25.9‰ in the 13C-PS treated limnocorral (Table 1). Within individual collection times, the greatest difference in both δ13C-DIC and δ13C-DOC between the treatment and control values was 2.1‰ (Fig. 1). There were no statistically significant differences in the slope or intercept of the relationship between δ13C and time for DIC (test for homogeneity of slopes, F3,8 = 0.823, p = 0.517, Fig. 1) or DOC (test for homogeneity of slopes, F3,8 = 0.491, p = 0.699, Fig. 1).
Fig. 1.
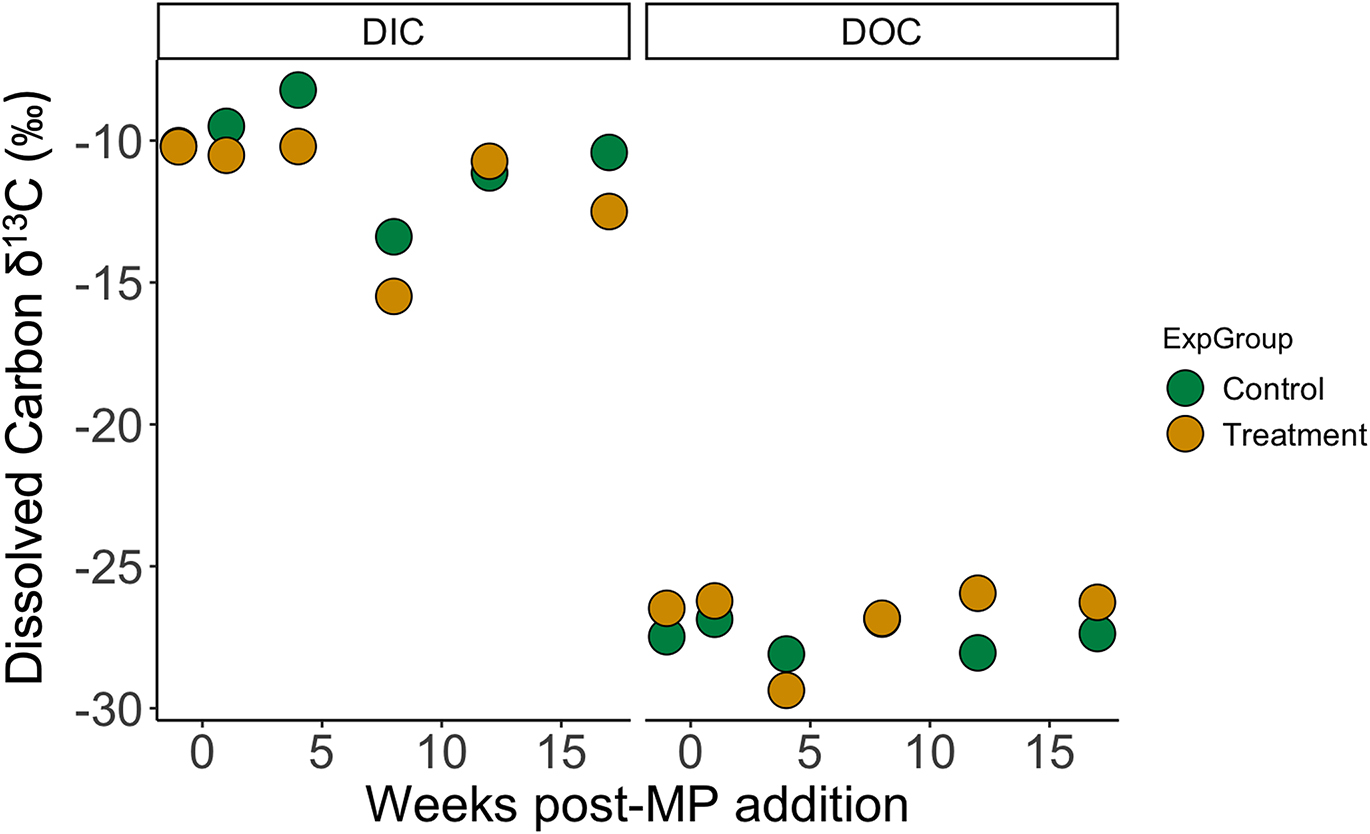
Based on the amount of 13C added to the limnocorral as microplastics, and the measured amount of C in the limnocorrals present as DOC, we estimated that DOC leached from microplastics at a rate of approximately 0.69% year−1 and, based on this leaching rate, the total mass of DOC leached from plastic over the 4-month experiment was 0.11 mg (see calculations in Supporting Information, Part D).
Compound specific δ13C in amino acids
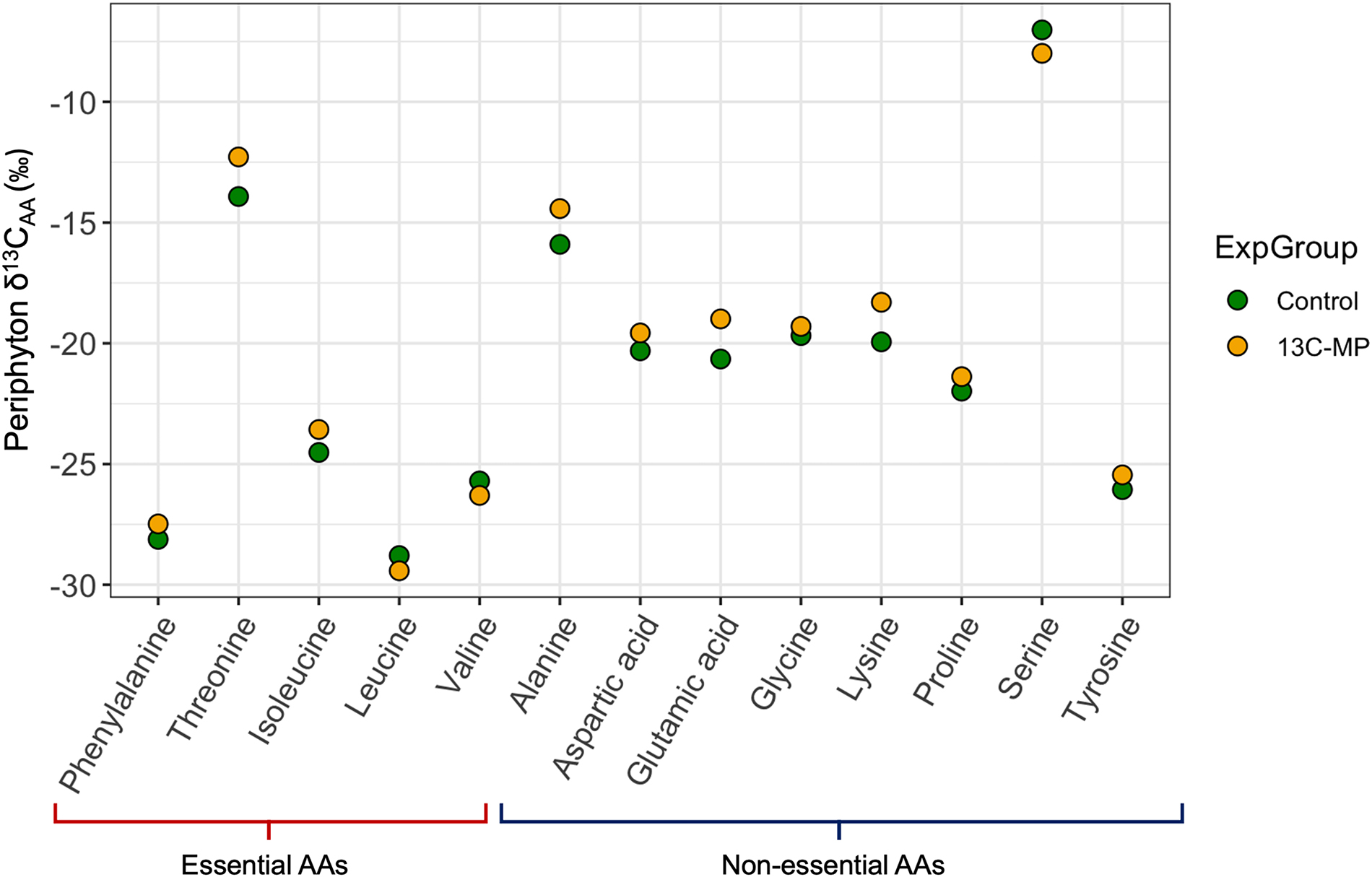
We confirmed that δ13CAA values of plankton (Fig. 2) and periphyton (Fig. 3) were most similar to that of bacteria and microalgae by comparison with Larsen et al. (2013; Table S2). We observed no changes in δ13C of amino acids that were greater than the range in natural variability expected for these biota (Figs. 2 and 3; see Larsen et al. 2013 for comparison of biota δ13CAA values). The largest difference between the 13C-PS treated limnocorral and the control was 1.7‰ for Threonine in plankton (Fig. 2), and 1.66‰ for Glutamic acid in periphyton (Fig. 3). Although the differences in δ13CAA between control and treatment samples were small (∼0.2 to 2‰; Figs. 2 and 3), the δ13CAA values were significantly greater in the 13C-PS treated limnocorral relative to the control for both plankton (t test, t = 2.036, p = 0.031, Fig. 2) and periphyton (t test, t = 1.810, p = 0.047, Fig. 3). For plankton, 11 out of the 14 measured amino acids were enriched in 13C and the mean difference between control and treatment δ13CAA was 0.50 ± 0.91‰. Similarly, for periphyton, 10 out of the 14 measured amino acids were enriched in 13C and the mean difference between control and treatment δ13CAA was 0.49 ± 1.01‰ (Fig. 3).
Fig. 2.
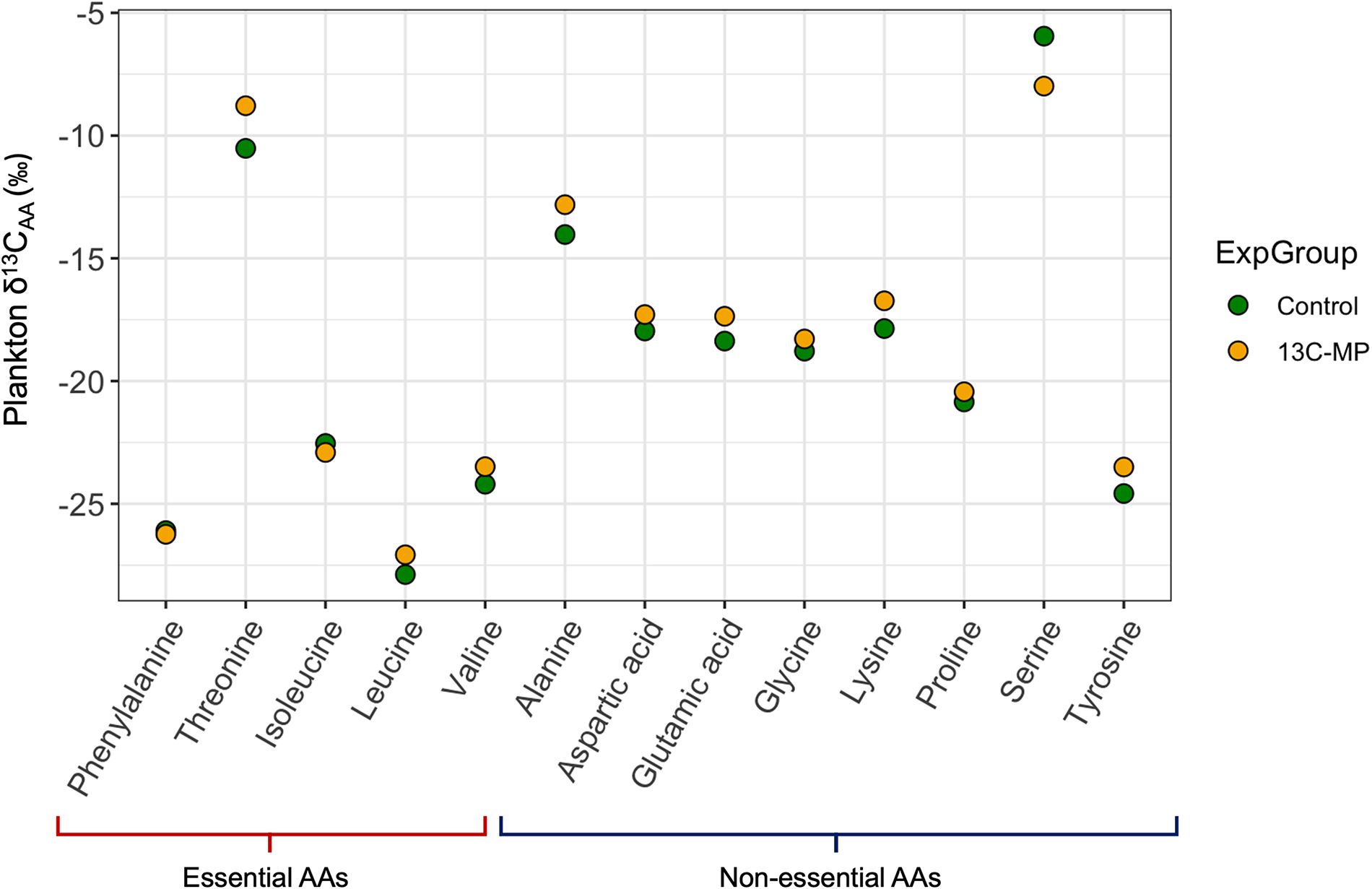
Fig. 3.
Discussion
We investigated the degradation of microplastics under natural conditions using 13C-PS in a limnocorral experiment. The results of this small study highlight that microplastic degradation is a slow process under natural conditions, and more work is needed to understand the timescale of degradation in the environment. We found minimal evidence of microplastic degradation and C leaching as DIC, DOC, and in periphyton and plankton. The value of this work, as well as its limitations and opportunities for future research, are discussed below.
Based on the relative difference in δ13C-DOC between the control and treatment limnocorral, we estimated that the leaching rate of plastic C herein was 0.69% year−1. Due to this slow rate of microplastic degradation, we were unable to detect any significant DOC production using δ13C-DOC in the present experiment outside the range of natural variability. Since this experiment was completed, two laboratory studies measuring degradation rates of 13C-labelled microplastics (polyethylene (PE) and PS) in humic and clear lake water have been conducted. In closed bottle experiments with organic C-depleted natural lake water, 13C-labelled PS was added and δ13C was measured in gas and microbial fatty acids over 6 weeks (Vesamaki et al. 2022). Polystyrene degraded at a rate of 0.22% year−1 in humic lake water and 0.17% year−1 in clear water (Vesamaki et al. 2022). Estimated degradation rates of PE in similar bottle experiments, again conducted with humic and clear lake water, were 0.45% year−1 in humic lakes and 0.07% year−1 in clear lakes (Taipale et al. 2022). Despite the use of different polymers (PE vs. PS), with different chemical and physical properties (PE is an aliphatic, positively buoyant polymer vs. PS is an aromatic, neutrally buoyant polymer), their calculated rates of degradation in humic lakes were similar to that measured herein (0.69% year−1).
Potential differences in environmental conditions between these laboratory studies versus the field (i.e., differences in UV radiation, temperature, microbial community composition) did not appear to largely influence the degradation rate of PS, but both studies highlighted that microbial community composition will strongly influence the biodegradation rates of microplastics because certain species are capable of decomposing plastic polymers (Taipale et al. 2022; Vesamaki et al. 2022). In particular, species in humic lakes may be more adept at decomposing strongly recalcitrant materials like plastic, although this appeared to be more important for PE than for PS. The lake studied herein was not particularly humic (DOC = 7.27–9.15 mg/L), although we did not study microbial community composition in the limnocorrals and cannot confirm whether any known species capable of biodegrading microplastics were present.
Using the degradation rate of PS calculated by Vesamaki et al. (2022), we re-evaluated the changes in δ13C we would expect to see over our experimental period. At a degradation rate of 0.22% year−1 we estimated that δ13C-DOC herein would increase 0.4‰ relative to the control value (see calculations in Supporting Information, Part C). Indeed, we did observe a change of this magnitude for PS in the 13C-microplastic limnocorral and calculated a similar degradation rate of 0.69% year−1 using our δ13C-DOC data (see calculations in Supporting Information, Part D), but this rate was not statistically significant based on our experimental design. Based on the low variability in control values throughout the experiment, and these low degradation rates estimated previously, our inability to detect changes in δ13C of DOC and DIC were likely related to low degradation rates and low mineralization of DOC and DIC in the water column rather than limited statistical power.
At the time this experiment was conducted, only one previous study, to our knowledge, had reported rates of microplastic degradation in terms of C loss. When plastic C loss and DOC production of expanded PS (EPS) was measured during a 2-month photochemical degradation experiments, total plastic C loss was 7% over the 2 months, and total DOC production was 6.8% of initial plastic C (Zhu et al. 2020). We initially used this laboratory-derived rate of EPS degradation to estimate the amount of C that could be expected to leach from the 13C-PS following addition to the limnocorral in Lake 378. Based on their findings, we calculated that if 7% of added 13C-PS leached as DOC over 2 months, measured δ13C-DOC in the experimental limnocorral would be 7.4‰, a value well outside the range of control δ13C-DOC values herein and the natural range of –29 to –19‰ (see Supporting Information, Part A: Table S1 and Zigah et al. 2011). However, degradation of C from microplastics in the natural environment we investigated herein was much slower than this previous laboratory experiment would suggest, and δ13C-DOC in our experimental limnocorral never exceeded –25.9‰, indicating that degradation of plastic could not be detected under the experimental conditions. While a slower degradation rate of PS herein could be expected given that PS has since been shown to leach C at a slower rate than EPS (Romera-Castillo et al. 2022), comparisons of our current experiment to Zhu et al. (2020) are largely limited because of differences in UV radiation, which often initiates the breakdown of microplastic polymers (Emad and Raghad 2013; Chamas et al. 2020). The present study was conducted outdoors under natural UV conditions in a lake, while Zhu et al. (2020) irradiated microplastics under a solar simulator, which could have a substantial influence on the measured degradation rate of microplastic and result in the contrasting degradation rates observed. Another difference between our study and Zhu et al. (2020) was the use of freshwater versus seawater, but this apparently does not have an effect on C leaching from microplastics (Egea et al. 2024).
Our study of the incorporation of plastic-derived C into the food web is comparable to another laboratory experiment by Taipale et al. (2023), wherein microplastic PS degradation and C incorporation was measured and compared to terrestrial leaves and lignin, similarly small amounts of 13C labelled PS were measured in zooplankton. Considerably larger amounts of 13C-PS (30 mg) were added to smaller volumes of lake water (2.6 L), but still, only small amounts of 13C-PS were detected in amino acids (up to 3%) and fatty acids (up to 2.5%; Taipale et al. 2023). Similarly, herein, most amino acids were significantly (albeit only slightly) enriched in 13C in the treatment limnocorral relative to the control (0.2–1.7‰ in 11 out of 14 AAs for plankton, 10 out of 14 AAs for periphyton). Collectively, these results may suggest evidence of degradation of microplastics at a similar rate. Unfortunately, we cannot definitively state whether our small increases in δ13C-DOC and most of the δ13CAA values were a result of a small amount of leached 13C-microplastic or simply natural variability in these endpoints due to limitations in the design of this pilot-scale in situ experiment.
In the present study, if we had anticipated a much slower degradation rate and added a greater amount of 13C-PS, we may have been able to detect greater 13C enrichment. For instance, we estimated based on our observed leaching rate (0.69% year−1) we would need to add 430 mg of 13C-PS to detect δ13C-DOC outside the range of natural variability (i.e., >–19‰). We also estimated that based on this leaching rate, in the current study it would have taken 2.9 y to detect δ13C-DOC outside the range of natural variability (i.e., >–19‰). In addition, a longer experimental period may have helped to detect 13C enrichment; however, we ran the experiment as long as possible during the full open-water season (May–September) and were limited by destruction of the mesocosms by ice-off in the spring. Further, due to the logistical difficulty of recollecting and counting these small microplastic particles, we cannot confirm the physical fate of the microplastics. We designed the experiment anticipating that the majority of PS fragments of this size range would remain in suspension for the 4-month experiment, in agreement with residence time calculations for PS done by Elagami et al. (2022). However, in contrast to previous literature and the known properties of virgin PS (Driedger et al. 2015; Rochman et al. 2019), companion studies in Lake 378 revealed that neutrally buoyant polystyrene either collected in a microlayer on the water surface or sank rapidly from water to sediment in limnocorrals in this lake (Rochman et al. 2024; Graves et al. Pers. Comm.). Therefore, any MP-derived C would more likely be water-surface or sediment-associated than in the water column. In retrospect and in future studies, given this new information, sampling could focus on the water surface and/or sediment and pore-water to understand the fate of MP-derived C.
In combination with slow degradation rates, one possible reason that we could not detect any significant plastic-derived C in the water column is that these leached compounds are expected to be highly biolabile and may be readily assimilated or utilized by microbial communities (Romera-Castillo et al. 2018; Zhu et al. 2020). Further, Taipale et al. (2019) also observed that C from plastic was more likely respired than utilized in macromolecules, suggesting that it may be more difficult to detect leached C incorporated into amino acids. Our experiment took place outdoors in the natural environment using 1 m wide in-lake enclosures that were open to the sediment and atmosphere, whereas previous studies have been conducted using bottle experiments in the laboratory (Taipale et al. 2019, 2022; Zhu et al. 2020). We likely encountered greater background noise and more difficulty detecting significant changes in 13C. For instance, we were not able to detect even a slight enrichment of 13C in DIC in the present experiment, suggesting that either respired DIC derived from microplastics was present only in a low/nondetectable amount, or potentially this produced gas quickly evaded to the atmosphere and was not accurately captured in our aqueous DIC samples.
Conclusion
This experiment was, to our knowledge, the first to study the ultimate fate of MP-derived C in a natural aquatic environment using 13C-labelled plastic. Over the 4-month experiment, only slight enrichments of 13C in DOC and some amino acids were observed relative to the control, and all measured stable isotope values were within the range of natural values for δ13C-DIC, δ13C-DOC, and δ13C-AAs, making it difficult to discern between natural variability and the slow degradation of microplastics. These results indicate very small amounts of C derived from added microplastics was found in the water column as DIC, DOC, and amino acids, suggesting that microplastic degradation is a slow process that could not be detected using 13C labelled plastic at this concentration (3268 particles/L) in the natural environment.
Ultimately, we observed minimal evidence of MP-derived C being incorporated into the aquatic food web in a boreal lake at the time scale we were able to examine these processes. We observed only small enrichments of 13C in DOC and some amino acids relative to the control, but these measurements were all within the range of natural variability for these endpoints and therefore cannot be attributed to microplastic degradation. Our results highlight that while microplastics can break down and components taken up by organisms in the lab, these processes are likely significantly slower in natural settings. This highlights the persistence of these plastics—even very small microplastics—to degradation processes that occur naturally, meaning that plastics persist for a very long time in the environment. These results highlight the need to perform more environmentally relevant, field-based studies to understand the fate and degradation of microplastics in the environment and will complement the other mesocosm and whole-lake experiments being conducted at the IISD-ELA to understand the fate and effects of microplastics in the lake environment (Rochman et al. in review). We note the importance of conducting environmentally realistic experiments to understand microplastic degradation and fate, as environmental conditions will influence the weathering and degradation of microplastics in ways that cannot be predicted or mimicked in the laboratory.
Acknowledgements
This is Contribution No. 6 of the pELAstic Project (https://thepelasticproject.com/). We thank Michael Paterson and Matthew Hoffmann for co-leading the pELAstic Project with CR, DO, MR, and JP. We are grateful to the International Institute for Sustainable Development for the opportunity to conduct experimental manipulations at the ELA field station, and their support of the pELAstic Project.
References
Bade D.L., Carpenter S.R., Cole J.J., Hanson P.C., Hesslein R.H. 2004. Controls of δ13C-DIC in lakes: geochemistry, lake metabolism, and morphometry. Limnology and Oceanography, 49: 1160–1172.
Bucci K., Tulio M., Rochman C.M. 2020. What is known and unknown about the effects of plastic pollution: a meta-analysis and systematic review. Ecological Applications, 30.
Chamas A., Moon H., Zheng J., Qiu Y., Tabassum T., et al. 2020. Degradation rates of plastics in the environment. ACS Sustainable Chemistry & Engineering, 8: 3494–3511.
Chen S.M., Mudie P., Sherwood O.A. 2022. Amino acid δ13C and δ15N fingerprinting of sea ice and pelagic algae in Canadian Arctic and Subarctic Seas. Frontiers in Marine Science, 9: 976908.
Curran K., Strlič M. 2015. Polymers and volatiles: using VOC analysis for the conservation of plastic and rubber objects. Studies in Conservation, 60: 1–14.
Dees J.P., Ateia M., Sanchez D.L. 2021. Microplastics and their degradation products in surface waters: a missing piece of the global carbon cycle puzzle. Environmental Science Technology Water, 1: 214–216.
Dreidger A.G.J., Durr H.H., Mitchell K., Van Cappellen P. 2015. Plastic debris in the Laurentian Great Lakes: a review. Journal of Great Lakes Research, 41: 9–19.
Egea L.G., Brun F.G., Jimenez-Ramos R. 2024. Dissolved organic carbon leaching from microplastics and bioavailability in coastal ecosystems. Science of the Total Environment, 909: 168673.
Elagami H., Ahmadi P., Fleckenstein J.H., Frei S., Obst M., Agarwal S., Gilfedder B.S. 2022. Measurement of microplastic settling velocities and implications for residence times in thermally stratified lakes. Limnology and Oceanography, 67: 934–945.
Emad Y., Raghad H. 2013. Photodegradation and photostabilization of polymers, especially polystyrene: review. SpringerPlus, 2: 398.
Graves S.D., Liber K., Palace V., Hecker M., Doig L.E., Janz D.M. 2021. Trophic dynamics of selenium in a boreal lake food web. Environmental Pollution, 280: 116956.
Horton A., Walton A., Spurgeon D.J., Lahive E., Svendsen C. 2017. microplastics in freshwater and terrestrial environments: evaluating the current understanding to identify the knowledge gaps and future research priorities. Science of the Total Environment, 586: 127–141.
Larsen T., Ventura M., Andersen N., O'Brien D.M., Piatkowski U., McCarthy M.D. 2013. Tracing carbon sources through aquatic and terrestrial food webs using amino acid stable isotope fingerprinting. PLoS ONE, 8: e73441.
Lee Y.K., Hong S., Jun H. 2021. A fluorescence indicator for source discrimination between microplastic-derived dissolved organic matter and aquatic natural organic matter. Water Research, 207: 117833.
Liu Y., Li J., Parakhonskiy B.V., Hoogenboom R., Skirtach A., De Neve S. 2024. Labelling of micro- and nanoplastics for environmental studies: state-of-the-art and future challenges. Journal of Hazardous Materials, 462: 132785.
McIlwraith H.K., Dias M., Orihel D.M., Rennie M.D., Harrison A.L., Hoffman M.J., et al. 2024. A multicompartment assessment of microplastic contamination in semi-remote boreal Lakes. Environmental Toxicology and Chemistry, 43: 999–1011, (in press).
Orihel D.M., Paterson M.J., Gilmour C.C., Bodaly R.A., Blanchfield P.J., Hintelmann H., et al. 2006. Effect of loading rate on the fate of mercury in littoral mesocosms. Environmental Science & Technology, 40: 5992–6000.
Posit team 2024. RStudio: Integrated Development Environment for R. Posit Software, PBC. Boston, MA. Available from http://www.posit.co/.
Priya K.L., Renjith K.R., Joseph C.J., Indu M.S., Srinivas R., Haddout S. 2022. Fate, transport and degradation pathway of microplastics in aquatic environment—a critical review. Regional Studies in Marine Sciences, 56: 102647.
Rochman C., Bucci K., Langenfeld D., McNamee R., Veneruzzo C., Covernton G., et al. 2024. Informing the exposure landscape: The fate of microplastics in a large pelagic in-lake mesocosm experiment. Environmental Science & Technology, 58: 7988–8088.
Rochman C.M., Brookson C., Bikker J., Djuric N., Earn A., et al. 2019. Rethinking microplastics as a diverse contaminant suite.Environmental Toxicology and Chemistry, 38: 703–711.
Romera-Castillo C., Birnstiel S., Álvarez-Salgado X.A., Sebastián M. 2022. Aged Plastic Leaching of Dissolved Organic Matter Is Two Orders of Magnitude Higher Than Virgin Plastic Leading to a Strong Uplift in Marine Microbial Activity. Frontiers in Marine Science, 9.
Romera-Castillo C., Pinto M., Langer T.M., Alvarez-Salgado X.A., Herndl G.J. 2018. Dissolved organic carbon leaching from plastics stimulates microbial activity in the ocean. Nature Communications, 9: 1430.
Sander M., Kohler H.P.E., McNeill K. 2019. Assessing the environmental transformation of nanoplastic through 13C-labelled polymers. Nature Nanotechnology, 14: 301–303.
Sheridan E.A., Fonvielle J.A., Cottingham S., Zhang Y., Dittmar T., Aldridge D.C., Tanentzap A.J. 2022. Plastic pollution fosters more microbial growth in lakes than natural organic matter. Nature Communications, 13: 4175.
Silfer J.A., Engel M.H., Macko S.A., Jumeau E.J., 1991. Stable carbon isotope analysis of amino acid enantiomers by conventional isotope ratio mass spectrometry and combined gas chromatography/isotope ratio mass spectrometry. Analytical Chemistry, 63(4): 370–374.
St. Jean G. 2003. Automated quantitative and isotopic (13C) analysis of dissolved inorganic carbon and organic carbon in continuous flow using a total organic carbon analyser. Rapid Communications in Mass Spectrometry, 17: 419–428.
Stainton M.P., Capel M.J., Armstrong F.A.J. 1977 The chemical analysis of freshwater, 2nd ed. Can Fish Mar Serv Misc Spec Publ. 25.
Taipale S.J., Peltomaa E., Kukkonen J.V.K., Kainz M.J., Kautonen P., Tiirola M. 2019. Tracing the fate of microplastic carbon in the aquatic food web by compound-specific isotope analysis. Scientific Reports, 9: 19894.
Taipale S.J., Rigaud C., Calderini M.L., Kainz M.J., Pilecky M., Uusi-Heikkila S., et al. 2023. The second life of terrestrial and plastic carbon as nutritionally valuable food for aquatic consumers. Ecology Letters, 26: 1336–1347.
Taipale S.J., Vesamaki J., Kautonen P., Kukkonen J.V.K., Biasi C., Nissinen R., Tiirola M. 2022. Biodegradation of microplastic in freshwaters: a long-lasting process affected by the lake microbiome. Environmental Microbiology, 1–12.
Vesamäki J.S., Nissinen R., Kainz M.J., Pilecky M., Tiirola M., Sami J. 2022. Decomposition rate and biochemical fate of carbon from natural polymers and microplastics in boreal lakes. Frontiers in Microbiology, 13: 1041242.
Wang W., Gao H., Jin S., Li R., Na G. 2019. The ecotoxicological effects of microplastics on aquatic food web, from primary producer to human: a review. Ecotoxicology and Environmental Safety, 173: 110–117.
Zhu L., Zhao S., Bittar T.B., Stubbins A., Li D. 2020. Photochemical dissolution of buoyant microplastics to dissolved organic carbon: rates and microbial impacts. Journal of Hazardous Materials, 383: 121065.
Zigah P.K., Minor E.C., Werne J.P., McCallister S.L. 2011. Radiocarbon and stable carbon isotopic insights into provenance and cycling of carbon in Lake Superior. Limnology and Oceanography, 56: 867–886.
Supplementary material
Supplementary Material 1 (DOCX / 3.12 MB).
- Download
- 3.12 MB
Supplementary Material 2 (XLSX / 14 KB).
- Download
- 13.23 KB
Information & Authors
Information
Published In

FACETS
Volume 10 • 2025
Pages: 1 - 9
Editor: Britt Dianne Hall
History
Received: 5 June 2024
Accepted: 1 December 2024
Version of record online: 14 February 2025
Copyright
© 2025 The Authors. This work is licensed under a Creative Commons Attribution 4.0 International License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Data Availability Statement
The data presented in this manuscript are available on the Borealis pELAstic project data repository (https://borealisdata.ca/dataverse/pELAstic).
Key Words
Sections
Subjects
Authors
Author Contributions
Conceptualization: SG, BH, CMR, JFP, MDR, DLM, ML, DMO
Data curation: SG
Formal analysis: SG, SMC, OAS
Funding acquisition: JFP, DMO
Investigation: SG, RM, THR, CMR, DLM, ML
Methodology: SG, SMC, BH, CMR, JFP, DLM, ML, OAS, DMO
Project administration: SG, DMO
Resources: CMR, JFP, MDR, DLM, ML, OAS, DMO
Supervision: BH, CMR, JFP, DMO
Visualization: SG
Writing – original draft: SG
Writing – review & editing: SMC, RM, THR, BH, CMR, JFP, MDR, DLM, ML, OAS, DMO
Competing Interests
The authors declare there are no competing interests.
Funding Information
Natural Sciences and Engineering Research Council of Canada: 401208952, RGPIN-2020-06956
Environment and Climate Change Canada: 3000760428
Mitacs: IT25640
This project was financially supported by an Environment and Climate Change Canada contract (3000760428 to DO) and a Natural Sciences and Engineering Research Council of Canada (NSERC) Discovery Grant (RGPIN-2020-06956 to DO). SG was supported by a MITACS Accelerate PDF Fellowship (IT25640, co-supervised by DO and BH) and a NSERC CREATE Scholarship (401208952, supervised by DO). We also acknowledge funding from Loblaws (to CMR) for supporting station fees, and ECCC (JP) for purchase of the 13C polystyrene used in this experiment.
Metrics & Citations
Metrics
Other Metrics
Citations
Cite As
Stephanie Graves, Shao-Min Chen, Rachel McNamee, Tazi H. Rodrigues, Brian Hayden, Chelsea M. Rochman, Jennifer F. Provencher, Michael D. Rennie, Daniel Layton-Matthews, Matthew Leybourne, Owen A. Sherwood, and Diane M. Orihel. 2025. Investigating the potential uptake of microplastic-derived carbon into a boreal lake food web using carbon-13 labelled plastic. FACETS.
10: 1-9.
https://doi.org/10.1139/facets-2024-0108
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
There are no citations for this item